

LONG-TERM SPATIO-TEMPORAL DYNAMICS OF SESSILE BENTHOS IN A SHALLOW MARINE SUBMERGED CAVE IN THE WESTERN MEDITERRANEAN SEA
Abstract
Submarine caves are considered priority habitats according to the Barcelona Convention and the EU Habitat Directive, yet they have received relatively low research effort when compared to other Mediterranean coastal habitats. This paper provides information about thelong-term spatio-temporal observations of sessile assemblages along the exterior-interior axis of Cerro-Gordo marine cave (Granada, Spain) from 2007 to 2016. Using feeding strategies and morphological descriptors, significant temporal variations of sessile community were recorded in both external and internal sectors, in spite of the higher confinement of the inner parts of the cave. In the inner cave sector, the use of feeding strategies revealed community variability through time and between opposite cave walls. On the other hand, the use of morphological descriptors proved to better depict variability in the outer cave sector, where community was more heterogeneous. These results provide valuable information that could be included in future monitoring programs of these endangered and protected habitats.
Key-words: Marine Cave, Sessile Benthos, Morphological Descriptor, Feeding Strategies
Introduction
Submarine caves are considered as key habitats and are included in the European Community Habitat Directive 92/43 EEC (habitat type 8330) and the “Dark Habitats Action Plan” (UNEP-MAP-RAC/SPA, 2015) for the conservation of sensitive species and habitats. They are important biodiversity hotspots which harbour rich biodiversity in coastal rocky areas (Gerovasileiou & Voultsiadou, 2012). In addition, these unique ecosystems are vulnerable and act as ecological islands, hosting communities sensitive to fragmentation as well as relict species and communities which are often similar to those found in abyssal systems (Boury-Esnault et al., 1993). The physicochemical conditions within submerged marine caves are easily disturbed by natural and anthropic alterations at global (e.g. climate change or water acidification) and regional/local scales (e.g. storms, coastal pollution, shoreline modifications or diving activities - Chevaldonné & Lejeusne, 2003; Parravicini et al., 2010; Di Franco et al., 2010). These alterations may affect benthic community at functional level (i.e. species morphology and feeding strategies -Guarnieri et al., 2012; Nepote et al., 2017; Montefalcone et al., 2018).
Given the variety of source of impacts and biotic responses, as well as the natural spatial heterogeneity and inaccessibility of these habitats, the effectiveness of efficient monitoring programs is often limited by the scarcity of information concerning the natural temporal changes present in these complex and variable systems (Magurran et al., 2010). Medium/long-term time series observations (years-decades) are essential tools in order to address these issues. The present study investigates the benthic community variability within a nine-year period in a shallow blind-ending submerged cave of the Western Mediterranean Sea (Cerro-Gordo cave, Alboran Sea).
Materials and methods
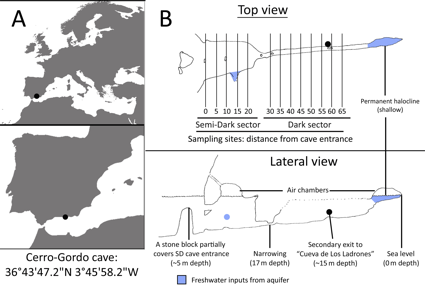
The cave of Cerro-Gordo is located in the North Coast of the Alboran Sea, within the limits of Maro & Cerro-Gordo Natural Park, in Granada, Spain (Fig. 1A).Benthic community was studied in the outer Semi-Dark (SD) and inner Dark (D) sectors of the cave (Fig. 1B). Within each sector, four 20 x 20 cm quadrats were randomly photographed at 5 m intervals on both walls of the cave, at a depth range of 6-8 m. Sampling took place in August of the years: 2007, 2010 and 2016 (D) and 2008, 2011 and 2016 (SD).
 |
 |
Benthic community coverage was obtained by overlaying 50 random points on each photoquadrat using photoQuad software (Trygonis & Sini, 2012). All taxa were assigned to two functional descriptors: Morphological descriptor (Md) and Feeding Strategies (FS), whose indicators were selected considering previous studies (Parravicini et al., 2010; Guarnieri et al., 2012; Gerovasileiou et al., 2017; Nepote et al., 2017 -see indicators on Results section). Orthogonal and fixed factors Year (three levels: 2007/2008, 2010/2011 and 2016) and Wall (two levels: Left and Right Wall, looking from the entrance of the cave) were tested separately for SD and D sectors. For each sector, multivariate PERMANOVA and SIMPER analyses were carried out using Md and FS datasets. Statistical analyses were carried out using PRIMER-E v6 + PERMANOVA (Andersonet al., 2008; Clarke & Gorley, 2006).
Results
Mean biotic coverage was similar in both Semi-Dark (SD: 62.40%) and Dark (D: 61.94%) sectors. However, the coverage of the used Md and FS variables differed between the two sectors (Fig. 2A and 3A).
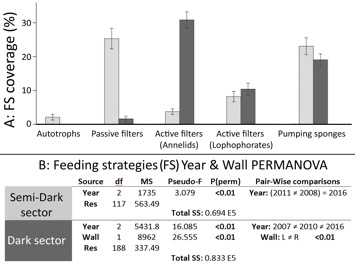
Temporal variation occurred in both cave sectors when grouping species coverage by feeding strategies (see Fig. 2B). In SD, the increase of passive filters (cnidarians) and the decrease of active pumping sponges contributed up to 67% of the accumulated dissimilarities between 2008 and 2011 (SIMPER). In D sector, lophophorates were responsible for 42% of multivariate dissimilarity between walls (higher coverage on the right wall) and active filter feeders grouped up to 60% of dissimilarities among years (temporal increase of lophophorates and annelids).
When grouping species coverage by morphologies, up to 78% of dissimilarities among years in SD sector (see Fig. 3B) were provoked by variations of prostrate sheets, flattened mounds and hemispherical mounds (SIMPER results). On the other hand, prostrate sheets, which were ubiquitous in the inner cave sector (Fig. 2A) contributed up to 80% of dissimilarities among years and walls in D.
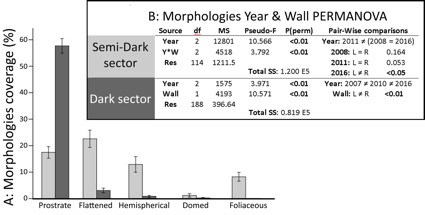
Morphological descriptor (Md) pooled higher variation than feeding strategies (FS) in outer (SD) sector, while both FS and Md pooled similar variation in D sector (see total sums of squares –SS– in Figs 2B and 3B). Note that FS and Md are different ways of grouping the same data, which allow comparisons by the amount of variation grouped by each descriptor (SS) and also by the distribution of this variation among factors and residuals (mean square sums –MS). See Andersonet al.(2005; 2008) for more information about this topic.
Discussion
Benthic community patterns with the use of Feeding Strategies (FS) and Morphological (Md) descriptorswithin Cerro Gordo cave were similar to those described for other Mediterranean marine submerged caves(see Rastorguef et al., 2015 and references therein). The predominance of prostrate life-forms in the inner parts of the cave has been suggested as an adaptation to a resource-restricted environment (Okamura et al.,2001). The presence of fine sediment in the bottom of D sector (see Navarro-Barranco et al., 2012) suggests that the tunnel connecting “Cerro-Gordo” to “Cueva de los Ladrones” does not significantly increase water motion. Yet, temporal variation in Cerro-Gordo cave benthic community was relatively similar in the outer and inner sectors, which could indicate the presence of water flow because of this secondary opening.
Cerro-Gordo cave was found to experience greater temporal changes than those usually expected for isolated ecosystems such as submerged marine caves. It is possible that frequent SCUBA diving activities within both cave sectors could have produced punctual turbidity events which may cause sponge mortalities (see Di Franco et al., 2010 and Nepote et al.,2017), and resuspension of large-size particles which may favour lophophorates and annelids(Riisgård & Larsen, 2000; Mancenido & Gourvennec, 2007). High-temperature peaks and storms in the study area became more frequent from 2012 onwards (Puertos del Estado, 2018), possibly influencing community functional dynamics. The deleterious effect of thermal rise in Mediterranean marine caves affects mainly massive sponges (Chevaldonné & Lejeusne, 2003; Parravicini et al., 2010; Montefalcone et al.,2018). It is also possible that D sector receives organic matter from percolation through small rock cracks and fissures. Such inputs may have a significant effect in highly oligotrophic ecosystems such as caves (Rastorgueff et al., 2015 and references therein).
In the outer SD sector, high non-linear temporal variation in the coverage of the anthozoans Astroides calycularis and Parazoanthus axinellae were the main contributors to the patterns observed. This may have modified cave community functioning and affected epibenthic assemblages (Martíet al.,2004; Navarro-Barranco et al.,2014; 2016). Finally, spatial differences within D sector for the cave wall communities are probably related to their inclination. The latter allows higher sediment deposition over left wall surfaces, leading to a reduction of Novocrania anomala brachiopod assemblages, which are known to inhabit mud-free surfaces (Rosso et al.,2015).
The results of this study support the effectiveness of both FS and Md descriptors for monitoring marine caves, as suggested by previous studies (Guarnieri et al., 2012; Nepote et al., 2017; Montefalcone et al., 2018). Nevertheless, morphological descriptors were more efficient for identifying variation inthe outer cave sector while the use of feeding strategies was more suitablein the inner cave sector where prostrate sheet forms dominate.
Conclusions
The spatio-temporal variation in Cerro-Gordo cave reflects complex context- and scale-dependent processes in marine caves with complex topographies (Harmelin 1969; Bussotti et al., 2006; Radolovićet al., 2015;Montefalcone et al., 2018). Sessile benthic community in Cerro-Gordo cave was found to be sensitive to external perturbations in accordance to studies in other Mediterranean marine caves (Chevaldonné & Lejeusne, 2003; Parravicini et al., 2010; Rastorgueff et al., 2015). These results provide valuable information for future monitoring programs for these fragile protected habitats.
Bibliography
ANDERSON M.J.,DIEBEL C.E., BLOM W.M., T.J. LANDERS (2005) - Consistency and variation in kelp holdfast assemblages: spatial patterns of biodiversity for the major phyla at different taxonomic resolutions. J. Exp. Mar. Bio. Ecol., 320: 35–56.
ANDERSON M.J., GORLEYR.N., CLARKEK.R. (2008) - PERMANOVA + for PRIMER. Guide to software and statistical methods.Plymouth, UK. PRIMER-E Ltd.
BOURY-ESNAULT N., HARMELIN J.G., VACELET J. (1993) - Les abysses méditerranéennes à vingt mètres de profondeur. La recherche,24: 848-851.
BUSSOTTI S., TERLIZZI A., FRASCHETTI S., BELMONTE G., BOERO F. (2006) - Spatial and temporal variability of sessile benthos in shallow Mediterranean marine caves. Mar. Ecol. Prog. Ser.,325: 109-119.
CHEVALDONNÉ P., LEJEUSNE C. (2003) - Regional warming-induced species shift in north-west Mediterranean marine caves. Ecol. Lett.,6: 371-379.
CLARKE K.R., GORLEY R.N. (2006) - PRIMER v6: User manual/tutorial. Plymouth: PRIMER-E
DI FRANCO A., FERRUZZA G., BAIATA P., CHEMELLO R., MILAZZO M. (2010) - Can recreational scuba divers alter natural gross sedimentation rate? A case study from a Mediterranean deep cave. ICES J. Mar. Sci.,67: 871-874.
GEROVASILEIOU V., VOULTSIADOU E. (2012) - Marine Caves of the Mediterranean Sea: A Sponge Biodiversity Reservoir within a Biodiversity Hotspot. PloS ONE,7: e39873.
GEROVASILEIOU V., DIMITRIADIS C., ARVANITIDIS C., VOULTSIADOU E. (2017) - Taxonomic and functional surrogates of sessile benthic diversity in Mediterranean marine caves. PloS ONE,12: e0183707.
GUARNIERI G., TERLIZZI A., BEVILACQUA S., FRASCHETTI S. (2012) - Increasing heterogeneity of sensitive assemblages as a consequence of human impact in submarine caves. Mar. Biol.,159: 1155-1164.
HARMELIN J.G. (1969) - Bryozoaires des grottes sous-marines obscures de la région marseillaise, faunistique et écologie. Téthys, 1: 793–806.
MAGURRAN A.E., BAILLIE S.R., BUCKLAND S.T., DICK J.M., ELSTON D.A., SCOTT E.M., SMITH R.I., SOMERFIELD P.J., WATT A.D. (2010) - Long-term datasets in biodiversity research and monitoring: assessing change in ecological communities through time. Trends Ecol. Evolut.,25: 574-582.
MANCENIDO M.O., GOURVENNEC R. (2007) A reappraisal of feeding current systems inferred for spire‐bearing brachiopods. Earth Env. Sci. T. R. So., 98: 345–356.
MARTÍ R., URIZ M.J., BALLESTEROS E., TURON X. (2004) - Temporal variation of several structure descriptors in animal-dominated benthic communities in two Mediterranean caves. J. Mar. Biol. Assoc. U. K., 84: 573-580.
MONTEFALCONE M., DE FALCO G., NEPOTE E., CANESSA M., BERTOLINO M., BAVESTRELLO G., MORRI C., NIKE BIANCHI C. (2018) - Thirty year ecosystem trajectories in a submerged marine cave under changing pressure regime. Mar. Environ. Res.,137: 98-110.
NAVARRO-BARRANCO C., GUERRA-GARCÍA J.M., SÁNCHEZ-TOCINO L., GARCÍA-GÓMEZ J.C. (2012) - Soft-bottom crustacean assemblages in Mediterranean marine caves: the cave of Cerro Gordo (Granada, Spain) as case study. Helgol. Mar. Res.,66: 292.
NAVARRO-BARRANCO C., GUERRA-GARCÍA J.M., SÁNCHEZ TOCINO L., GARCÍA-GÓMEZ J.C. (2014) - Mobile epifaunal community in marine caves in comparison to open habitats. Aquatic Biol.,20: 101-109.
NAVARRO-BARRANCO C., GUERRA-GARCÍA J.M., SÁNCHEZ-TOCINO L., FLORIDO M., GARCÍA-GÓMEZ J.C. (2016) - Amphipod community associated with invertebrate hosts in a Mediterranean marine cave. Mar. Biodivers.,46: 105-112.
NEPOTE E., BIANCHI C.N., MORRI C., FERRARI M., MONTEFALCONE M. (2017) - Impact of a harbour construction on the benthic community of two shallow marine caves. Marine Poll. Bull., 114: 35-45.
OKAMURA B., HARMELIN J.G., JACKSON J.B.C. (2001) - Refuges revisited: enemies versus flow and feeding as determinants of sessile animal distribution and form. In: Jackson J.B.C., Lidgard S., Mckinney F.K. (Eds.) Evolutionary Patterns: Growth, Form and Tempo in the Fossil Record. University of Chicago Press, Chicago: 61–93 pp.
PARRAVICINI V., GUIDETTI P., MORRI C., MONTEFALCONE M., DONATO M., BIANCHI C.N. (2010) - Consequences of sea water temperature anomalies on a Mediterranean submarine cave ecosystem. Estuar. Coast. Shelf Sci., 86: 276-282.
PUERTOS DEL ESTADO (2018) - Prediccion de oleaje, nivel del mar; Boyas y mareografos. Datos históricos, SIMAR: 2042080 and SIMAR: Málaga [online] Available at: http://www.puertos.es/es-es/oceanografia/Paginas/portus.aspxMinisterio de Fomento, Gobierno de España. [last accessed on october 2018]
RADOLOVIĆ M., BAKRAN-PETRICIOLI T., PETRICIOLI D., SURIĆ M., PERICA D. (2015) - Biological response to geochemical and hydrological processes in a shallow submarine cave. Mediterr. Mar. Sci.,16: 305-324.
RASTORGUEFF P.A., BELLAN-SANTINI D., BIANCHI C.N., BUSSOTTI S., CHEVALDONNÉ P., GUIDETTI P., RUITTON S. (2015) - An ecosystem-based approach to evaluate the ecological quality of Mediterranean undersea caves. Ecol. Indic.,54: 137-152.
RIISGÅRD H.U., LARSEN P. S. (2000) - Comparative ecophysiology of active zoobenthic filter feeding, essence of current knowledge. J. Sea Res.,44: 169–193.
ROSSO A., SANFILIPPO R., MATROTOTARO F. (2015) - Bryozoan and serpulid distribution pattern on deep/water slab (Bari Canyon, Adriatic Sea). In: UNEP/MAP - RAC/SPA.Proceedings of the 1st Mediterranean Symposium on the conservation of Dark Habitats, Slovenia. 79-80 pp.
TRYGONIS V., SINI M. (2012) - photoQuad: a dedicated seabed image processing software, and a comparative error analysis of four photoquadrat methods.J. Exp. Mar. Biol. Ecol.,424: 99-108.
UNEP-MAP-RAC/SPA (2015). - Action Plan for the Conservation of Habitat and Species Associated with Seamounts, Underwater Caves and Canyons, Aphotic Hard Beds and Chemo-syntetic Phenomena in the Mediterrranean Sea. Dark Habitats Action Plan. RAC/SPA, Tunis: 17 pp
Juan SEMPERE-VALVERDE, SABINO L. A., ESPINOSA F., GEROVASILEIOU V., SÁNCHEZ-TOCINO L., NAVARRO-BARRANCO C.
Laboratorio de Biología Marina (LBM), Facultad de Biología, Universidad de Sevilla, Avda. Reina Mercedes s/n, Campus de Reina Mercedes, 41012, Sevilla, Spain.
E-mail: juansempere91@gmail.com