

La concha de los moluscos es una estructura de una importancia adaptativa esencial para la defensa de animal y, subsidiariamente, como anclaje y soporte de las partes blandas. Se trata de un carácter ya presente en los moluscos más ancestrales, aparecidos hace unos 540 m.a, y quizás antes (sobre 555 m.a.). Algunos moluscos han reducido o perdido la concha secundariamente y la han sustituido por defensas químicas (nudibranquios) o por una gran eficiencia hidrodinámica (cefalópodos endococleados).
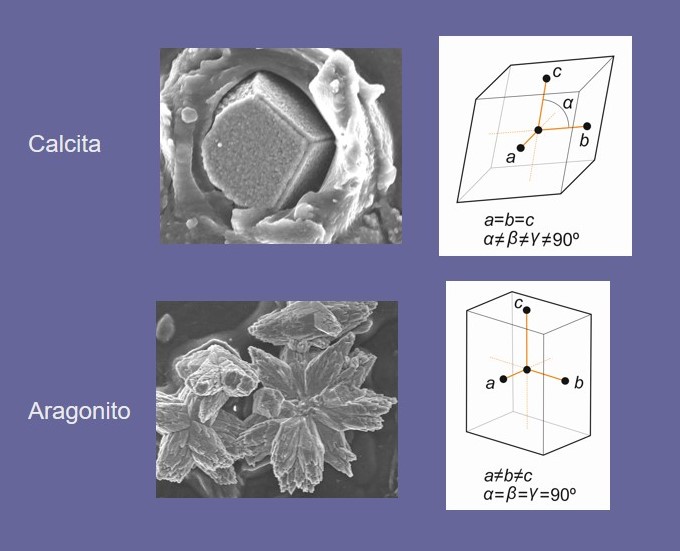
La concha de los moluscos está formada por dos minerales, aragonito y calcita, cuya composición es la misma: carbonato cálcico. De ambos minerales se dice que son polimorfos de carbonato cálcico y la diferencia entre ambos es la ordenación de sus moléculas dentro de redes cristalinas con diferente simetría, lo que se denomina sistemas cristalinos (trigonal en calcita y ortorrómbico en aragonito) (Fig. 1). Estas diferencias provocan que los cristales de ambos minerales tengan morfologías, y propiedades mecánicas y ópticas diferentes.
Cuando se observa la fractura de una concha, incluso al ojo desnudo, inmediatamente se aprecia que se trata de un material cristalino. Dentro de las conchas, los cristales de calcita o aragonito presentan morfologías, orientaciones y relaciones mutuas recurrentes, denominadas microestructuras (Fig. 2). Las microestructuras aparecen en la mayoría de los invertebrados y unicelulares capaces de calcificar, pero es en moluscos donde aparece la mayor diversidad, seguidos a cierta distancia por los braquiópodos y los briozoos.
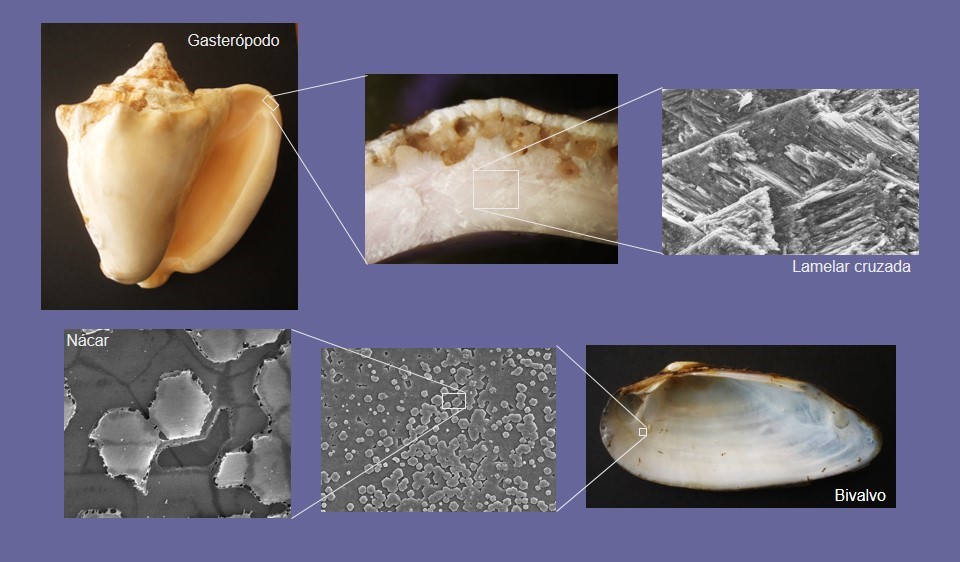
En moluscos se conocen unas 15 microestructuras básicas, aunque algunos autores diferencian hasta el doble de variedades microestructurales. Las más importantes son (Fig. 3):
Calcíticas:
- Prismática columnar: Prismas calcíticos, a veces de milímetros de longitud, perpendiculares a la superficie de la concha. Están muy bien individualizados a base de membranas orgánicas, de hasta varias micras de espesor. Propia de capas externas de bivalvos Ostreoida y algunos Pectinida (Propeamussiidae, Anomioidea).
- Prismáticas irregulares o granulares: Los prismas tienen contornos muy complicados y morfologías irregulares. Propia de las capas externas de bivalvos (algunos Anomalodesmata) y algunos gasterópodos.
- Fibrosa: Fibras paralelas y muy bien orientadas que forman un ángulo de unos 45º con la superficie de crecimiento. Propia de Mytilida.
- Foliada: Lajillas terminadas en punta de flecha, organizadas en láminas, que se inclinan a un ángulo pequeño con respecto la superficie interna de la concha. Muy característica de Pectinida y Ostreida, así como en algunos Patellogastropoda.
- Chalk: Es una microestructura a base de laminillas verticales, muy porosa (hasta un 80% en volumen de huecos) y se intercala, en forma de lentes dentro de las capas foliadas de Ostreidae.
- Lamelar cruzada: Consiste en paquetes perpendiculares a la superficie de la concha y que se alargan tanto en paralelo como en perpendicular al margen de la concha. Los paquetes contienen fibras de contorno rectangular que se inclinan entre 25° y 45° con respecto a la superficie de la concha. De modo característico, de fibras de paquetes consecutivos se inclinan en direcciones opuestas. Sólo aparece en Patellogastropoda.
Aragoníticas:
- Homogénea: Son granos de muy pequeño tamaño rodeados de finas membranas orgánicas. Se conoce en Nuculanida y algunos Heterodonta (ej., Glossus, Arctica).
- Fibrosa: Fibras finas de aragonito que radian a partir de un núcleo y que se dirigen perpendiculares o a un gran ángulo hacia la superficie de la concha. Aparece en las capas externas de Vetigastropoda y Heterodonta.
- Prismática: Existe una variante columnar que tiene las mismas características que su equivalente calcítica y en que el relleno de las unidades prismáticas es de aragonito fibroso (propia de Palaeoheterodonta, y algunos Monoplacophora), así como una variante granular consistente en prismas muy cortos embebidos en abundante matriz orgánica (aparece en algunos Anomalodesmata).
- Foliada: Igual que su equivalente calcítico, aunque las lajillas presentan terminaciones rectas o redondeadas. Sólo se conoce en algunos Monoplacophora.
- Nácar: Es, con mucho, la microestructura más conocida y estudiada, porque constituye las perlas de valor comercial. Consta de laminillas de contorno pseudohexagonal, rómbico, rectangular u ovalado que se empaquetan en láminas. Éstas se superponen de modo que los centros de las tabletas de las distintas láminas se alinean en la vertical (nácar columnar) o están mutuamente desplazados (en pared de ladrillo). Propia de Nuculida, Pterioidea, Pinnoidea, Vetigastropoda, Nautiloidea y algunos Monoplacophora.
- Lamelar cruzada: Igual que su equivalente calcítico. Es con mucho la microestructura más abundante en moluscos. Constituye las placas de Polyplacophora, las conchas de Scaphopoda, de la inmensa mayoría de Gastropoda (salvo algunos Vetigastropoda) y Bivalvia (Heterodonta, Arcida).
- Espiral: Fibras que forman espirales cuyos ejes son perpendiculares a la superficie de la concha, e idénticas en radio, inclinación y orientación. Sólo aparece en gasterópodos Cavolinioidea (Opisthobranchia).
- Prismas de contorno pseudohexagonal paralelos entre sí, que forman capas delgadas. Constituyen el material al que se anclan directamente los músculos de unión del cuerpo blando a la concha.

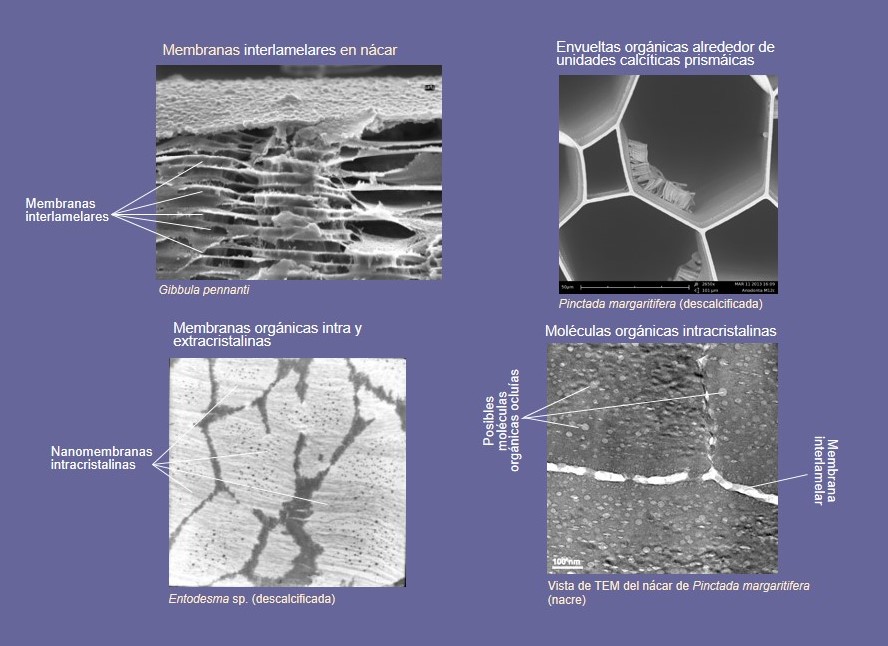
Junto con los cristales, existe siempre un cierto contenido en materia orgánica (1-10%), en forma de proteínas y péptidos, polisacáridos (β-quitina en moluscos) y lípidos. La mayor proporción dentro de la fracción orgánica corresponde al componente proteico. Un aspecto muy interesante de la materia orgánica es su distribución (Fig. 4). Una pequeña proporción queda atrapada en el interior de los cristales (materia orgánica intracristalina), tanto en forma de pequeñas oclusiones como de nanomembranas. Otra fracción forma membranas gruesas que pueden rodear las unidades de las capas prismáticas o formas finas membranas horizontales entre las lamelas de nácar. A pesar de su pequeña proporción, el papel del componente orgánico es esencial (véase más adelante) y es lo que marca la diferencia entre las microestructuras y los agregados cristalinos inorgánicos.
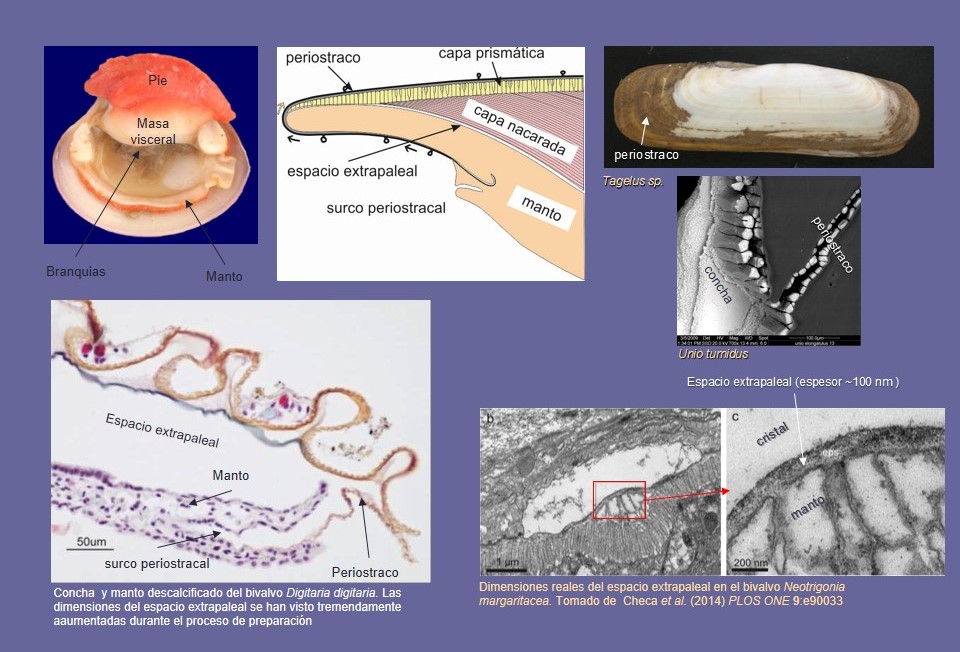
Las microestructuras (como la concha en general) están segregadas por el epitelio del manto de animal, que es una capa unicelular que reviste al manto y cuya superficie externa está directamente en contacto con la superficie interna (de crecimiento) de la concha (Fig. 5). Los mecanismos fisiológicos que tiene lugar durante la segregación de la concha son muy complejos y no son perfectamente conocidos. En breve, el manto se encarga de segregar al pequeño espacio que lo separa de la concha (denominado espacio extrapaleal; Fig. 5) los componentes necesarios para la fabricación de la concha: pequeños agregados de carbonato cálcico amorfo y compuestos orgánicos, que se unen posteriormente a la concha. El manto es un órgano extraordinario debido a su increíble capacidad de calcificar, ye que el peso de concha excede, en algunos casos, decenas de veces, al de las partes blandas.
Durante el mecanismo de segregación de la concha, la primera estructura que se segrega es una lámina orgánica llamada periostraco (Fig. 5). Se forma en un surco entre los lóbulos medio y externo del borde del manto y, durante el crecimiento, gira hacia el dorso y ocupa la posición más externa (Fig. 5). Una función obvia es cerrar el espacio extrapaleal y aislarlo del medio externo, con objeto de que los componentes necesarios para la formación de la concha no difundan al mismo. La segregación de la concha comienza en el mismo margen del manto por debajo del periostraco.
Comparado con los agregados cristalinos inorgánicos, las microestructuras de los moluscos y otros invertebrados son compuestos (se les denomina compuestos organo-minerales) especialmente sofisticados, en el sentido de que los cristalitos presentan morfologías muy diferentes de y ordenaciones mucho más elaboradas que las de equivalentes inorgánicos. En esta ordenación intervienen las moléculas orgánicas presentes, que determinan el tipo de polimorfo (aragonito, calcita), las morfologías cristalinas, y los mecanismos de nucleación y crecimiento. Además, en la organización de las microestructuras intervienen procesos de crecimiento cristalino, procesos físicos de autoorganización y procesos de actividad celular directa (reconocimiento por contacto, segregación local diferencial de productos orgánicos o minerales...). Todo ello hace que las microestructuras sean productos imposibles de reproducir en laboratorio con la tecnología actual, lo cual sería de gran interés en ciencia de materiales por sus elevadas capacidades biomécanicas (véase abajo).
Con el crecimiento del molusco, las microestructuras forman capas contínuas (salvo el chalk). Típicamente, la concha aumenta de espesor desde el dorso hacia el margen, aunque en algunos bivalvos ocurre lo contrario, bien porque las capas más internas se engruesen hacia el dorso, o porque se añadan nuevas capas en esta posición.
Las conchas tienen al menos dos capas (tres en bivalvos, si se cuenta la fina capa miostracal), usualmente con diferentes microestructuras, que, a veces presentan diferentes mineralogías (Fig. 6). En este caso, siempre son las capas calcíticas las que se colocan en posición externa. Un caso típico son los Pterioidea y Pinnoidea (Pteriomorphia), que constan de una capa externa prismática calcítica y otra capa interna aragonítica. A excepción de los gasterópodos Epitoniidae, no existen actualmente conchas totalmente calcíticas, ya que incluso las ostras y pectínidos presentan una capa miostracal (aragonítica). Contrariamente, sí existen grupos de concha enteramente aragonítica (la mayoría de los Caenogastropoda, los Protobranchia, Palaeoheterodonta, Arciida y la inmensa mayoría de Heterodonta). Dentro de Caenogastropoda, Patellogastropoda y Arcida se encuentran capas superpuestas con idéntica microestructura, siempre de tipo lamelar cruzada, cuya única diferencia es la orientación de las lamelas de primer orden, que pasan de ser comarginales a antimarginales, o viceversa, de una capa a otra (Fig. 6).
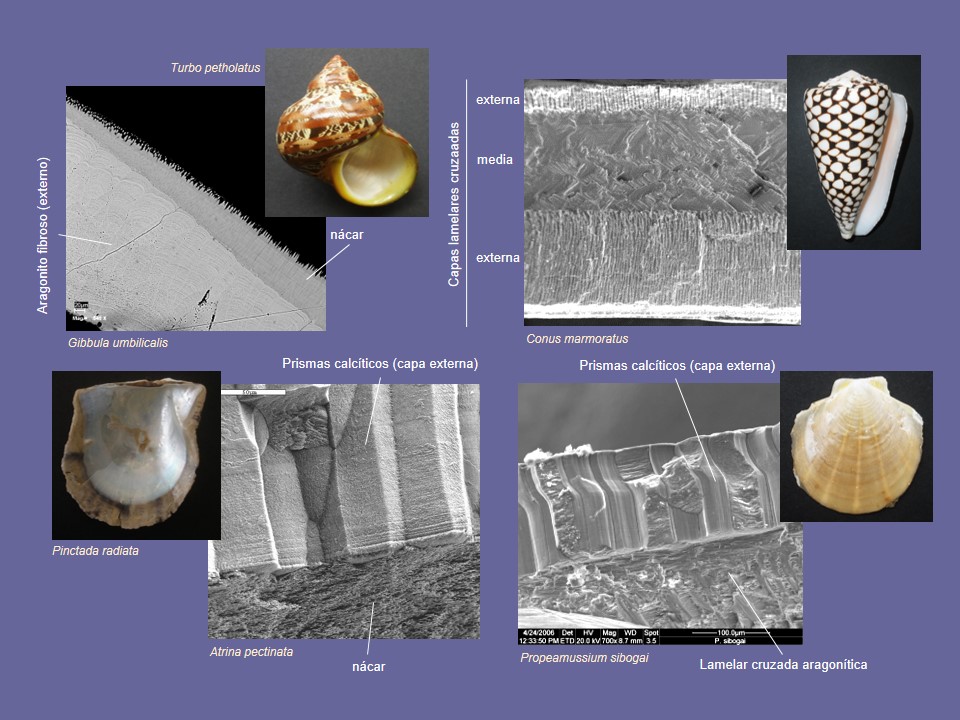
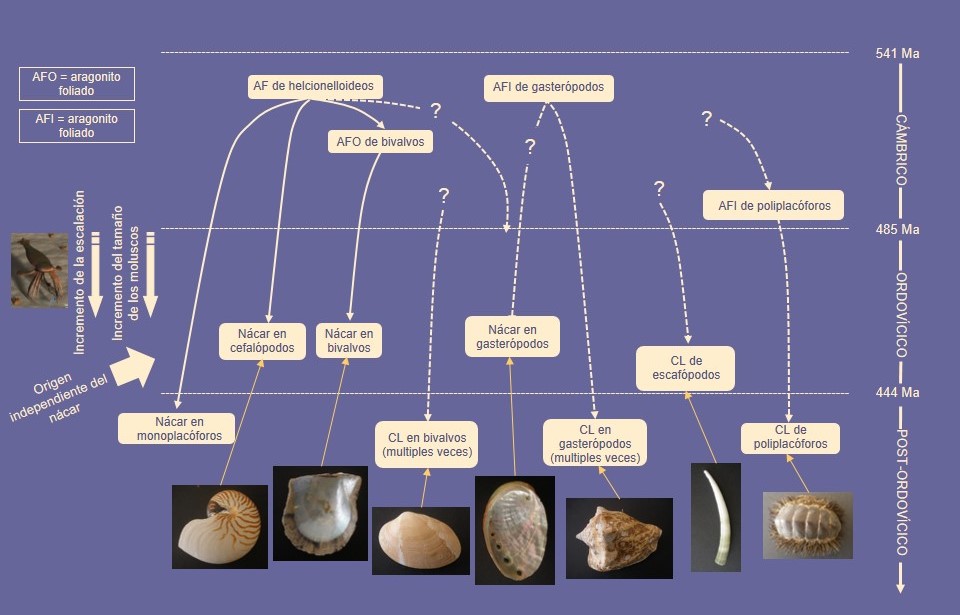
La microestructura es un carácter conservativo dentro de las superfamilias (Ostreoidea, Pinnoidea+Pterioidea), de los órdenes (Arcida, Limida, Mytilida, Pecinida, Nuculanida, Nuculida), de las subclases (Heterodonta, Palaeoheterodonta, Patellogastropoda, Caenogastropoda, Vetigastropoda) e incluso de las clases (Cephalopoda, Scaphopoda, Polyplacophora), y suele usarse como carácter útil en filogenia. No obstante, las mismas microestructuras en diferentes clases (y a veces dentro de ellas; ej. lamelar cruzada de bivalvos Arcida y Heterodonta) se han obtenido mediante convergencia (Fig. 7). El caso más llamativo es el del nácar, presente en bivalvos, gasterópodos, cefalópodos, y monoplacóforos, y que se ha desarrollado de modo independiente en las cuatro clases. Lo mismo puede decirse de la microestructura lamelar cruzada, segregada hoy en día por escafópodos, poliplacóforos, y gran cantidad de gasterópodos y bivalvos. Ello se sabe porque los ancestros comunes (ya extinguidos) de estos grupos y los representantes más basales de estas clases (aparecidos en el Cámbrico inferior) nunca segregaron ninguno de estos dos materiales, que aparecieron muy posteriormente (unos 60 m.a. después).
Las conchas cumplen básicamente una función de protección del animal frente a los depredadores. De hecho, están relativamente bien diseñadas para esta función, ya que, por ejemplo, con una rigidez parecida, su resistencia a la fractura es aproximadamente un orden y medio de magnitud mayor que la de los minerales correspondientes de carbonato cálcico inorgánicos (calcita, aragonito). La responsable de estas propiedades mecánicas superiores es la materia orgánica, no sólo por su cantidad, sino también por su distribución. Por ejemplo, de entre todas las microestructuras, la más resistente en tensión, compresión y flexión es el nácar, seguido de la microestructura lamelar-cruzada (Tabla 1).
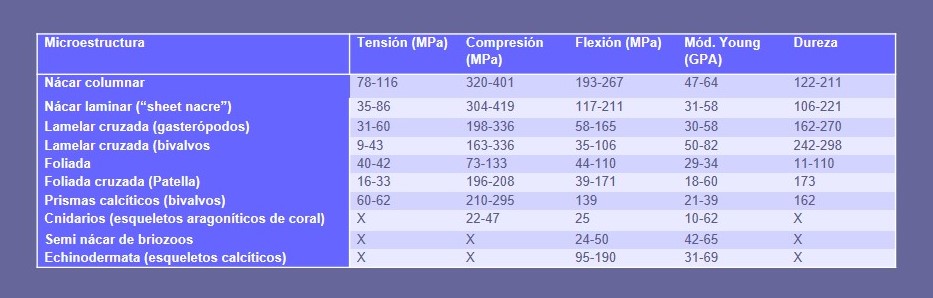
Las microestructuras calcíticas están muy por detrás en estos parámetros. El nácar está diseñado de modo que durante la deformación, las tabletas son capaces de deslizarse ligeramente a favor de las membranas interlamelares, y, cuando se produce la rotura, los planos siguen los contornos de las tabletas y de las membranas interlamelares. Ambos procesos contribuyen ampliamente a la disipación de energía. En el caso de la microestructura lamelar cruzada, los planos de fractura igualmente se desvían a lo largo de los contactos entre lamelas de diverso orden (Fig. 8).

Otras microestructuras están diseñadas para funciones distintas de la de resistencia. Por ejemplo, las microestructuras columnares calcíticas de Ostreida (Ostreoidea, Pinnoidea, Pterioidea) tienen una capacidad enorme para flexionarse, resultado de la existencia de fundas orgánicas deformables alrededor de las unidades prismáticas (Fig. 9).
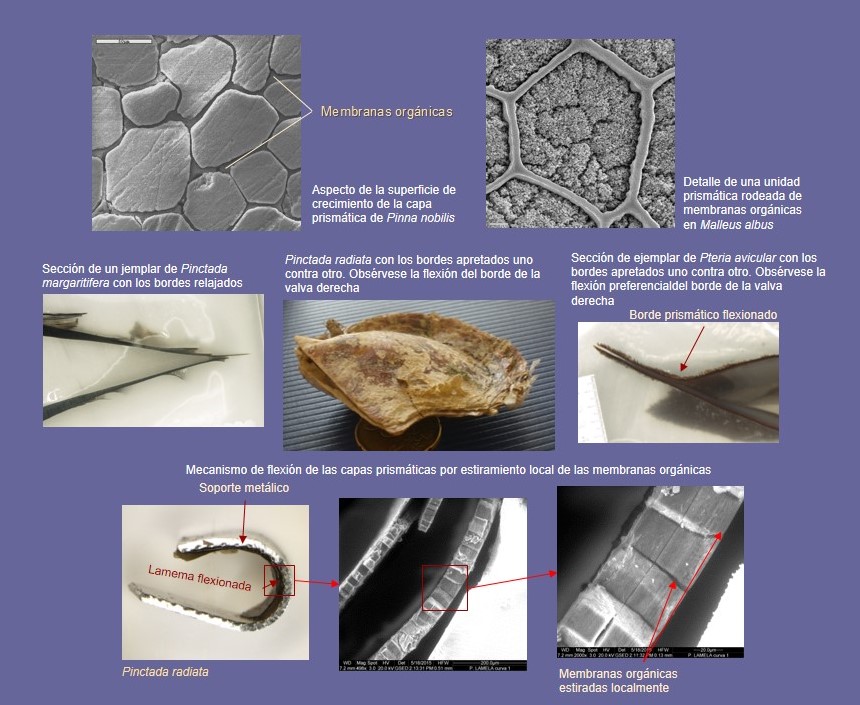
Durante el cierre de la concha, el margen de la valva derecha se dobla y se adapta al interior de la valva izquierda, que es más rígida. Así se consigue un cierre totalmente estanco de la concha, útil frente a depredadores, o en formas intermareales (como muchas ostras) como mecanismo para evitar la deshidratación. Las conchas calcíticas son sensiblemente más blandas que las aragoníticas, pero tienen la ventaja de que la fractura es localizada y no extensiva como en el caso de las conchas aragoníticas (Fig. 10). El chalk es un material muy blando y muy poroso, lo que permite segregar grandes espesores de concha ligera, con poco gasto energético y de material de construcción.

En este sentido es curioso que las microestructuras segregadas por los moluscos del Cámbrico (aragonito foliado y fibroso, semi-nácar calcítico) fuesen de muy escaso rendimiento biomecánico. No fue hasta el Ordovícico cuando apareció el nácar de modo independiente en gasterópodos, bivalvos y cefalópodos (en estos pudo aparecer en el Cámbrico superior), seguido algo después por las capas lamelares cruzadas. Este hecho es próximo en el tiempo al Gran Evento de Biodiversificación del Ordovícico, en que se produce además un incremento en la diversidad de depredadores (peces, cefalópodos, euriptéridos). Al mismo tiempo, los moluscos pasan de tener tallas milimétricas o submilimétricas a centimétricas, con lo que podrían haber quedado al alcance de estos predadores. El desarrollo de microestructuras de gran resistencia biomecánica podría haber sido la respuesta al incremento a la escalación propio de esta época (Fig. 7).
Con el tiempo, nácar y lamelar cruzada han ido siendo las microestructuras más abundantes, pero mientras que la primera alcanzó su máximo (en términos del número de familias que la segregaron) en el Paleozoico medio-superior, la microestructura lamelar cruzada ha seguido creciendo en importancia. Ello, en opinión de algunos autores, es consecuencia del alto coste energético de la fabricación del nácar (cuyo contenido en materia orgánica, la parte ‘costosa’, es de entre 4,5-5%), frente a la microestructura lamelar cruzada (que sólo contiene 1-1,5%). Pero, aparte, existe un componente filogenético claro, ya que aquellos grupos que han contribuído esencialmente a la diversificación de los moluscos (bivalvos Heterodonta y gasterópodos Caenogastropoda) tienen conchas exclusiva o casi exclusivamente a base de capas lamelares cruzadas.
ANTONIO CHECA GONZÁLEZ