

Reino: Animal; Filo: Chordata; Subfilo: Vertebrata; Superclase: Gnathostomata; Superclase: ? Tetrapoda; Clase: Mammalia; Subclase: ? Theria; Orden: Cetartiodactyla; Suborden: Cetancodonta; Infraorden: Cetacea
LOS CETÁCEOS
Los cetáceos comprenden un grupo amplio de mamíferos marinos. No todas las especies de mamíferos marinos se adaptan de igual forma al medio acuático. Es uno de los grupos que presenta el máximo nivel de adaptación a la vida en el agua, ya que son independientes del medio terrestre. Los apareamientos, nacimientos, alimentación y cualquier otro comportamiento tienen lugar en el agua. Algunas especies de cetáceos ocupan medios de agua dulce.
Delfines cazando peces voladores
“Cetáceo” es un término que agrupa a los animales que comúnmente conocemos como ballenas, delfines y marsopas. Existe gran diversidad entre las especies, no sólo en cuanto a formas y tamaños, sino también en conducta, hábitat, etc. Podemos encontrarlos en todos los mares de nuestro planeta, incluso hay especies que viven en los ríos. Este grupo abarca desde el animal más grande que existe en nuestro planeta, el rorcual azul, de más de 30 metros de longitud, hasta las pequeñas marsopas de poco más de un metro.

Las dimensiones de los cetáceos son muy variables. Oscilan entre 1,5 y 25 m en los odontocetos (cetáceos con dientes) y entre los 10 y 30 m en los misticetos (cetáceos con barbas). El tamaño relativo de los sexos es muy variable dentro de este grupo de cetáceos. Hay especies en las que los machos superan en tamaño a las hembras y otras en las que es al contrario.
Los odontocetos, generalmente tienen la mandíbula superior estrecha y recta, poseen al menos un par de dientes en la mandíbula inferior y un melón globoso por encima de los ojos. Sólo tienen una dentición a lo largo de su vida y son dientes homodontos. Algunas especies tienen decenas de dientes cónicos en cada lado de las mandíbulas mientras que otras tienen pocos y, algunas especies como el cachalote, los tiene sólo en la mandíbula inferior. Los dientes pueden ser variables en tamaño y forma y los usan para atrapar y sujetar a sus presas ya que no mastican el alimento.
La mandíbula superior de los misticetos es larga y sostiene una hilera de láminas córneas paralelas, flexibles y planas conocidas como “ballenas” o “barbas”, deshilachadas por el borde interior, que les permiten filtrar el alimento del agua. Las barbas varían en número, color y tamaño según la especie. Las mandíbulas inferiores carecen de barbas.
ADAPTACIONES MORFOLÓGICAS, ANATÓMICAS Y FISIÓLOGICAS DE LOS CETÁCEOS A LA VIDA ACUÁTICA
A lo largo de la evolución, el cuerpo de los cetáceos ha sufrido una serie de transformaciones morfológicas, anatómicas, ecológicas y de comportamiento, respecto a sus antecesores terrestres como adaptación a la vida en el mar.

Son mamíferos completamente acuáticos. El agua es mucho más densa (al menos tres veces más) y más viscosa (aproximadamente 60 veces mayor) que el aire a temperaturas similares. Ambas propiedades influyen en el rozamiento ya que son fuerzas que se oponen al movimiento de un cuerpo en un líquido, mucho más, si este es el medio marino, por lo que moverse en ella es más difícil y costoso. Además el agua absorbe más rápido el calor y la luz, aparte de transmitir el sonido a mayor velocidad y distancia. Otra propiedad es la presión, importante factor que influye en los organismos marinos, ya que a medida que aumenta la profundidad en la columna de agua, mayor es la fuerza que se ejerce sobre el cuerpo y lo comprime (esta fuerza es equivalente a 1 atmósfera por cada 10 metros de profundidad).
La forma del cuerpo de los cetáceos es de las mejores adaptadas para moverse ágil y rápidamente en el agua. Es muy hidrodinámica. Del cuerpo sólo sobresalen sus aletas, muy útiles para la natación. La cabeza es una de las partes de los cetáceos que mejor sirve para identificarlos. La mayoría presentan un rostro prominente terminado en pico, más o menos largo, por delante de los ojos.

El esqueleto de los cetáceos presenta numerosas modificaciones para adaptarse a la vida en el agua. El cráneo en comparación con sus antecesores terrestres se ha condensado y han sufrido un fenómeno conocido como “telescopización”. A lo largo de la evolución, los maxilares y premaxilares se fueron alargando hacia atrás, desplazando los huesos nasales hacia la zona dorsal. Como resultado la abertura nasal o espiráculo se encuentra en la parte dorsal del cuerpo, lo que constituye una adaptación a la vida acuática ya que les facilita mucho la respiración. Su forma y localización exactas varían según la especie. Los odontocetos presentan un espiráculo mientras que los misticetos presentan dos colocados en forma de V (Figura 1).
|
|
|


Una de las mejores formas de localizar cetáceos, sobre todo grandes cetáceos, en mar abierto es su soplo. El soplo es el resultado de la respiración del animal: una exhalación explosiva seguida inmediatamente de una inhalación, lo que produce una nube de gotitas de agua y vapor de agua sobre la cabeza del animal cuando respira. Los soplos varían en altura, forma y visibilidad según las especies y pueden llegar a ser muy distintos (Figura 2).
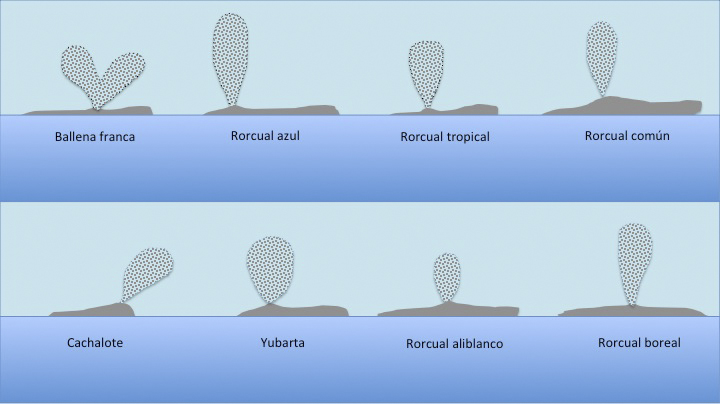
Este patrón de respiración para los cetáceos más pequeños sólo toma una fracción de segundo, pero puede durar unos segundos para las grandes ballenas. El soplo será más alto si se produce después de un buen tiempo de inmersión a cuando el animal se encuentra reposando en la superficie.
Soplo de un cachalote
El acto de respirar es consciente y voluntario. Debajo del agua no abren el espiráculo por lo que en caso de quedar atrapados morirían por asfixia y no ahogados. La laringe en los cetáceos está atravesada por el esófago. Esto determina que las vías respiratorias y digestivas sean independientes, de tal manera que los cetáceos pueden deglutir aunque estén sumergidos, sin que el agua pase nunca a las vías respiratorias (Figura 3)
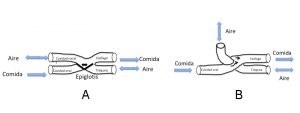
Los cetáceos odontocetos a pesar de tener dientes, no mastican la comida, directamente la tragan. El esófago es distendido y el estómago presenta generalmente tres cámaras aunque en algunas especies podrían ser más (Figura 4). Funcionalmente, estos compartimentos corresponden a las distintas regiones del estómago simple de la mayoría de los mamíferos. El intestino de los cetáceos es relativamente largo y no existe diferencia funcional entre intestino delgado y grueso en el caso de los odontocetos mientras que en misticetos es muy clara la diferencia entre intestino grueso y delgado. La parte inicial del mismo, el duodeno, se expande en un saco que ocasionalmente puede confundirse con un cuarto estómago.

Las vértebras cervicales, son aplanadas y en algunas especies podrían aparecer fusionadas algunas de ellas, por lo que la movilidad del cuello estaría muy restringida. Generalmente aparecen en número de siete. El número de vértebras torácicas, lumbares y caudales varía según la especie. Existen vestigios de los huesos de la cintura pélvica pero sin conexión con la columna vertebral.
La caja craneana se comprime y deja hueco para el melón. El melón es un órgano ovoide de naturaleza lipídica cuya función no ha sido del todo establecida, pero se cree que es un componente bioacústico usado en la ecolocalización, que consiste en la habilidad de analizar el ambiente circundante (detectando obstáculos, presas, predadores, etc.) emitiendo sonidos y escuchando el eco que éstos producen.
En los odontocetos las mandíbulas superior e inferior son muy variadas según la especie, pero casi siempre son rectas vistas de lado. Sólo tienen una dentición a lo largo de su vida con dientes homodontos. Algunos tienen decenas de dientes cónicos en cada lado de las mandíbulas mientras que otros tienen pocos y algunas especies como el cachalote, sólo en la mandíbula inferior. Los dientes pueden ser numerosos o reducirse a un solo par, como es el caso de los zifios. Los usan para atrapar y sujetar a sus presas ya que no mastican el alimento.
|
|
|


{kind=link}
{kind=link}
En los misticetos, la cabeza es aplanada y la mandíbula superior sostiene una hilera de láminas córneas flexibles y paralelas conocidas como “ballenas” o “barbas”, que corresponden a láminas de queratina con el borde interior deshilachado que les permiten filtrar el alimento del agua. Las barbas varían en número, color y tamaño según las especie (Figura 6). En las verdaderas ballenas las barbas son largas y la mandíbula está arqueada hacia arriba. Sin embargo, los rorcuales, las tiene cortas y la mandíbula es recta. Los misticetos son capaces de abarcar con su boca un gran volumen de agua, la cual expulsan después con la ayuda de la lengua atravesando las barbas. Así retienen a los pequeños animales de los que se alimentan, sobre todo crustáceos eufausiáceos (krill) que forman densos bancos y pequeños peces.
Debido a la gran flexibilidad que presenta la caja torácica es posible una extensión máxima de entrada de aire durante la inspiración a los pulmones y una comprensión máxima debida al aplastamiento por la presión durante las inmersiones profundas. En misticetos, su estructura anatómica es incapaz de soportar el peso del animal en caso de varamiento, por lo que suelen morir rápidamente, mientras que la de los odontocetos hace que soporten mejor su peso fuera del agua.
Las extremidades anteriores se han convertido en las conocidas como aletas pectorales, situadas por detrás de la cabeza y por debajo de la línea media del cuerpo, Todos los cetáceos tienen aletas pectorales bien desarrolladas, aunque pueden variar mucho en forma y tamaño según la especie (Figura 7).

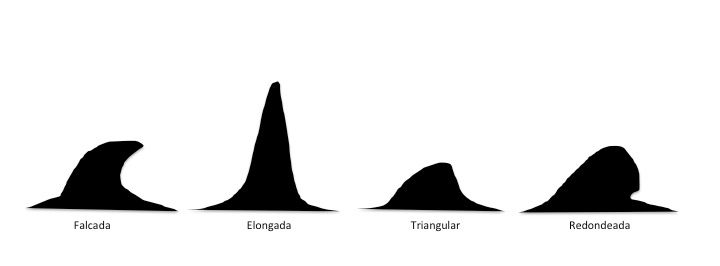
Tienen estructura ósea conectada a la columna vertebral y aunque mantienen la secuencia de huesos típica de otros muchos vertebrados, pueden variar en número y forma dependiendo de la especie. El número de dedos es variable (3, 4 ó 5) y generalmente presentan hiperfalangia. La articulación del codo es inmóvil mientras que la del hombro es bastante rígida, excepto en las yubartas. La función principal de estas aletas pectorales es estabilizadora y mantener el equilibrio además de permitirles girar. Sin embargo, las aletas dorsal y caudal no tienen hueso. Están formadas por tejido conectivo. La aleta dorsal varía en forma y tamaño en las diferentes especies (Figura 8), pudiendo incluso estar ausente en algunas, y facilita la termorregulación y la estabilidad dinámica.
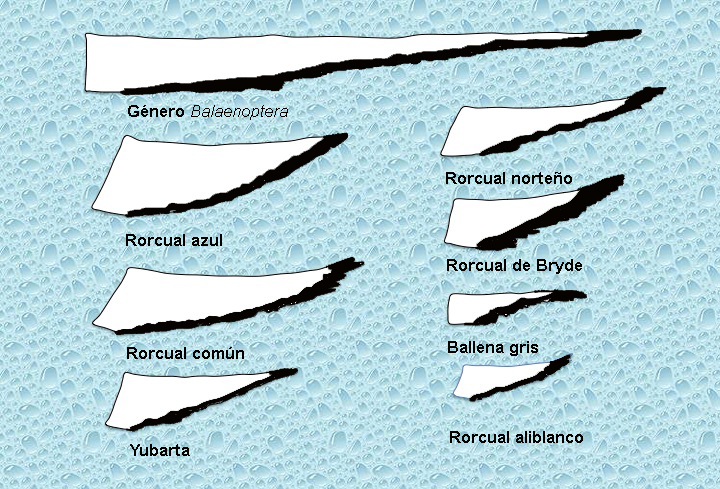
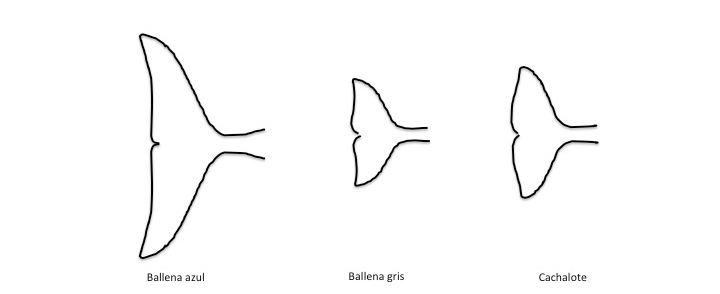
Las extremidades posteriores desaparecen y el cuerpo se estrecha en la parte posterior hacia el ano formando el rígido pedúnculo caudal que termina en la aleta caudal o cola. Dispuesta en un plano horizontal y está formada por tejido conectivo fibroso y elástico, es una aleta rígida a la vez que flexible que usan principalmente como elemento propulsor. Esta aleta también es variable en forma y tamaño según la especie (Figura 9).
Una adaptación muy importante de los cetáceos a la vida acuática es la Termorregulación. Estos animales han desarrollado una serie de estrategias para retener calor en ambientes fríos utilizando mecanismos fisiológicos, bioquímicos, anatómicos y etológicos. Para combatir la excesiva pérdida de calor se han provisto de una espesa capa de grasa que funciona como aislante térmico y como una importante reserva energética además de jugar un papel muy importante en los procesos de flotabilidad. Son animales endotermos ya que son capaces de mantener su temperatura corporal constante entre los 36 y 38 ºC independiente de la que haya en el medio exterior.
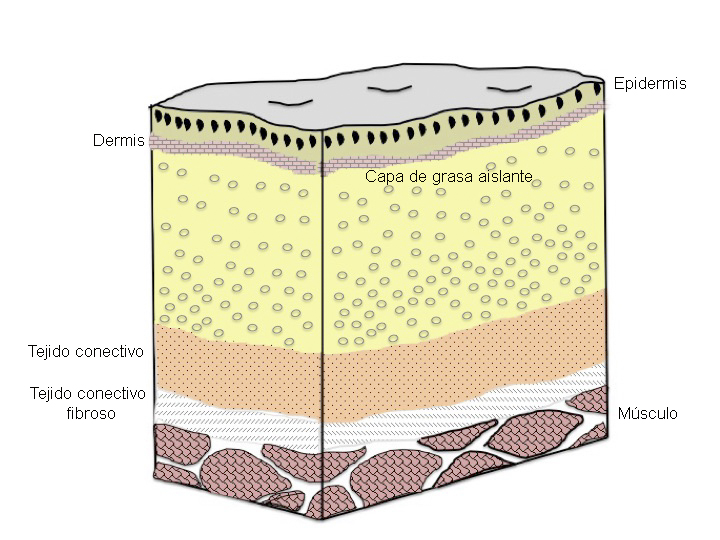
La piel de los cetáceos es lisa y tiene 3 capas, como en el resto de los mamíferos. La primera es la capa externa o epidermis, la segunda es la capa interna o dermis y la tercera es una capa de grasa que sirve de protección y para evitar las pérdidas de calor, llamada hipodermis. La capa de grasa en algunos cetáceos es especialmente gruesa, sobre todo en animales muy grandes, pudiendo sobrepasar los 50 cm de espesor (la capa de grasa de la ballena de Groenlandia (Balalena mysticetus) alcanza los 70 cm), mientras que en otros cetáceos es de sólo unos pocos centímetros. Esta capa de grasa o “blubber” contiene proteínas como el colágeno y la elastina así como numerosos vasos sanguíneos. Aunque generalmente representa el 30% del peso corporal del animal, su grosor varía dependiendo de la especie, de su ubicación geográfica, de las capacidades de buceo y del estado nutricional del animal. Debajo de la piel de las aletas no existe capa de grasa. El grosor de la piel varía a lo largo de la superficie corporal y con la edad del animal. Por debajo de la capa de grasa se encuentran los músculos (Figura 10).
Se sabe que los delfines mulares (Tursiops truncatus) renuevan o recambian la capa externa de la piel unas 12 veces al día (cada dos horas, nueve veces más rápido que en los humanos) lo que le permite mantener una piel lisa y suave que disminuya el rozamiento.
El suministro de sangre al cerebro en estos mamíferos marinos no tiene lugar por la vía normal de las arterias carótidas sino a partir de vasos asociados a la médula espinal que conforman la denominada “rete mirabile”. Consiste en una red compleja de venas y arterias muy próximas entre sí que utilizan un mecanismo de flujo a contracorriente y que juega un papel muy importante en los cetáceos en determinadas funciones como en la retención de agua y excreción de una orina hipertónica, en mantener a los testículos a una temperatura adecuada para su funcionamiento así como en el control de las pérdidas de calor a nivel de las aletas. El principio de contracorriente o contraflujo consiste en que el calor de la sangre que fluye hacia las aletas se transfiere a la sangre fría procedente de éstas, la cual se calienta antes de volver a fluir de nuevo hacia el resto del cuerpo (mientras que la sangre que fluye desde el cuerpo se va enfriando, lo que impide una pérdida excesiva de calor). Este curioso sistema puede controlar la cantidad de calor que se pierde en la superficie de las aletas; el calor se conserva cuando es necesario mediante el mecanismo de contracorriente o se disipa cuando este mecanismo está desconectado (si se produce un exceso de calor como resultado de un esfuerzo por ejemplo).

Entre las adaptaciones a los periodos de hipoxia (estado en el cual el cuerpo entero, hipoxia generalizada, o una región del mismo -hipoxia de tejido-, se ve privado del suministro adecuado de oxígeno) los pulmones están nada o poco lobulados, formando esencialmente dos lóbulos. Desde el punto de vista anatómico aparece un aumento de estructuras de soporte como pueden ser cartílago, colágeno, músculo liso y tejido elástico a nivel de los bronquiolos terminales, lo que facilita una apnea prolongada y un rápido intercambio de aire al emerger.
Otra adaptación a la hipoxia es que poseen un mayor volumen sanguíneo que el de los mamíferos de tamaño y peso similar (se sabe que los cetáceos poseen un número muy elevado de glóbulos rojos, de 7-11,5 mill/cc.) y una elevada concentración de hemoglobina en sangre y mioglobina en los músculos. Incluso que las células musculares poseen hemoglobina a la que se le denomina miohemoglobina. En cada inspiración son capaces de renovar entre el 80% y el 90% del aire de los pulmones frente al 10% o 20% de la mayoría de los mamíferos terrestres. Además, los cetáceos son capaces de restringir el riego sanguíneo sólo a órganos vitales durante una inmersión profunda, de manera que dichos órganos no se dañen por la falta de oxígeno.
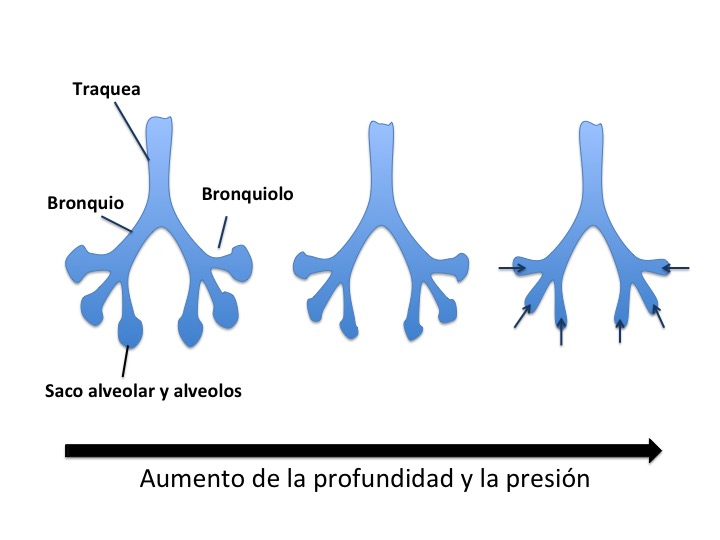
El oxígeno consumido por el metabolismo del cuerpo en una apnea (suspensión voluntaria de la respiración dentro del agua mientras se recorren largas distancias o se desciende hasta grandes profundidades) lo almacenan en tres compartimentos: el sistema respiratorio, la sangre y la musculatura corporal. Estos mamíferos marinos pueden hacer un uso eficaz del reservorio pulmonar de O2 durante buceos poco profundos, pero no son capaces de hacer lo mismo a grandes profundidades. Al comprimirse el aire en profundidad a medida que el animal se sumerge, los alvéolos serían la primera parte de los pulmones en colapsarse y de esta manera el aire quedaría en su mayor parte en las vías respiratorias aéreas de conducción (tráquea, bronquios y bronquiolos no respiratorios). Si ocurre esto, el O2 de los pulmones no quedaría disponible porque desde estas vías de conducción se transfiere muy poco oxígeno a la sangre. Ese colapso de los alveolos les sirve además para evitar la narcosis por nitrógeno, ya que durante el ascenso, evitarían la liberación del nitrógeno en forma de gas a la sangre, absorbido durante la inmersión. El colapso alveolar se produce gracias a una especie de esfínter que poseen en los bronquiolos terminales. Se cree que el colapso alveolar ocurre entre 30 y 100 m de profundidad.
Aunque muchos cetáceos cuentan con una gran capacidad visual, el oído es el órgano de los sentidos que se ha convertido cada vez más vital para los cetáceos, quienes además han afinado su destreza para la ecolocalización.
Los cetáceos tienen la vista bastante desarrollada. Para poder enfocar tanto dentro como fuera del agua, han adquirido una fuerte musculatura ocular que puede modificar la forma del cristalino para que se adapte a ambos tipos de visión. La acomodación del ojo se lleva a cabo por un mecanismo llamado “desplazamiento axial del cristalino” (se mueve de adelante hacia atrás) producido por cambios en la presión intraocular. El delfín mular es capaz de mover el ojo hacia fuera de la órbita unos 10-15 mm y retraerlo nuevamente. Este movimiento es muy común cuando observa binocularmente algún objeto fuera del agua. Otra habilidad que poseen es la capacidad de mover el ojo derecho independientemente del izquierdo y mantener un párpado abierto mientras el otro lo mantiene cerrado.
Pero la adaptación de la visión en los cetáceos también afecta al iris y a la pupila. Cuando el animal de desplaza hacia aguas profundas poco iluminadas, requiere que la pupila se dilate rápidamente y pase de una forma de U a tener forma esférica. Esta característica es común en muchos delfines y ballenas.

Otra adaptación que tienen los cetáceos a la poca luz es una capa reflectora bien desarrollada situada detrás del epitelio de la retina que produce múltiples reflexiones que inciden nuevamente en ella incrementando así la sensibilidad visual en condiciones de poca luminosidad. Esta capa se conoce como “tapetum lucidum” y está presente en todos los cetáceos. Hay evidencias considerables de la presencia tanto de conos como bastones en cetáceos pero sus retinas tienen dominancia de bastones con escasos conos (1% de fotoreceptores).
La piel de los cetáceos es lisa y carece casi por completo de pelo (aunque todas las especies tienen un manto de pelo durante el desarrollo fetal y algunos poseen algo de pelo en la zona facial en la vida adulta). Son pelos largos y sensitivos denominado vibrisas. En los odontocetos, sólo se pueden observar pelos, muy pocos, en el extremo superior del morro de los embriones y son poco numerosos en los misticetos.
El tacto en ellos tiene una gran importancia social intra e interespecífica, ya que son frecuentes los roces y el contacto sexual entre individuos, ya sean o no del mismo sexo y a veces entre individuos de distintas especies. Las mayores concentraciones de terminaciones nerviosas se localizan en las zonas más sensibles del cuerpo como son por ejemplo la cabeza y zonas próximas al espiráculo, a los ojos y a los genitales. La piel está provista de células sensibles que optimizan su destreza para nadar de manera rápida y eficiente y es muy sensible a estímulos de su medio ambiente.
La coloración típica de muchas especies cambia desde que nacen hasta que son adultos. Hay especies que nacen de un color más claro y se van oscureciendo al crecer y otras en las que ocurre a la inversa. Los misticetos presentan coloraciones más homogéneas que los odontocetos. En estos últimos, los patrones básicos de coloración de la piel suelen ser tonos de grises, con mayor claridad en el vientre y tonos más oscuros en el lomo. A menudo se combina con líneas y manchas de diferentes tintes y contrastes. Se pueden definir tres patrones básicos de coloración: patrón uniforme (un solo color ya sea claro u oscuro), patrón listado o moteado (con zonas fuertemente coloreadas sobre todo en la cabeza, aletas, costados y vientre) y Patrón contrasombreado (la parte dorsal del cuerpo siempre es de color oscuro y la parte ventral clara). Un ejemplo de cada uno de ellos serían la beluga, la orca y el delfín mular respectivamente. Generalmente los patrones de contrasombreado claro-oscuro conllevan una función principal de camuflaje y los patrones aposemáticos podrían estar relacionados con la comunicación intraespecífica, protección frente a depredadores, etc.

Audición y Producción de sonidos:
Los oídos de los cetáceos están situados justo detrás de los ojos y son pequeñas aberturas que carecen de pabellón auditivo externo. Su mandíbula inferior conduce muy bien los sonidos. En su interior, a lo largo de una cavidad que la recorre, contiene un tejido graso llamada “grasa mandibular”. Esta cavidad que contiene la grasa mandibular termina conectando con la placa timpánica. Hay evidencias de que utilizan su mandíbula inferior como un oído externo que recibe las ondas sonoras y a través de la grasa, los guía hasta el tímpano y de ahí al cerebro. En los misticetos, la ruta del sonido hacia el oído no está del todo clara ya que carecen tanto de canal mandibular como de la grasa en su interior.
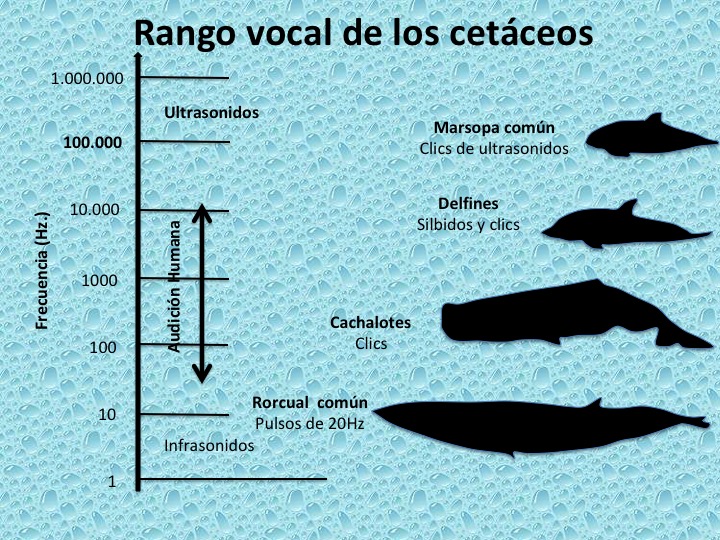
Los odontocetos son capaces de oír frecuencias muy altas (sonidos muy agudos de más de 150 KHz) y tienen una gran sensibilidad acústica en el agua (el rango de audición de un delfín mular es de 0,075-150 KHz, mientras que el de un humano es de 0,015-20 KHz). En contraste, los misticetos son capaces de producir sonidos de alto volumen en el rango infrasónico, de frecuencias inferiores a los 20 Hz (Figura 14).
Los odontocetos pueden producir tres tipos de sonidos: clicks, sonidos pulsados (chilllidos) y silbidos. No todos los odontocetos pueden producir todos estos tipos de sonidos. Los clicks, producidos por los sacos aéreos, se utilizan para los procesos de ecolocalización. Consisten en breves ráfagas de impulsos sonoros. Los delfines producen clicks de corta duración que se encuentran típicamente entre 40 y 300 kHz en la mayoría de las especies. Los silbidos y los sonidos pulsados (chillidos), son usados para la comunicación intraespecífica. Los silbidos son señales de frecuencia modulada de banda estrecha que varían desde cortas y simples a señales complejas y largas. Generalmente se sitúan entre 1-30 kHz. Los sonidos pulsados pueden ocurrir tan rápidamente, uno después de otro, que pueden ser percibidos como un sonido continuo (Watkins, 1967)
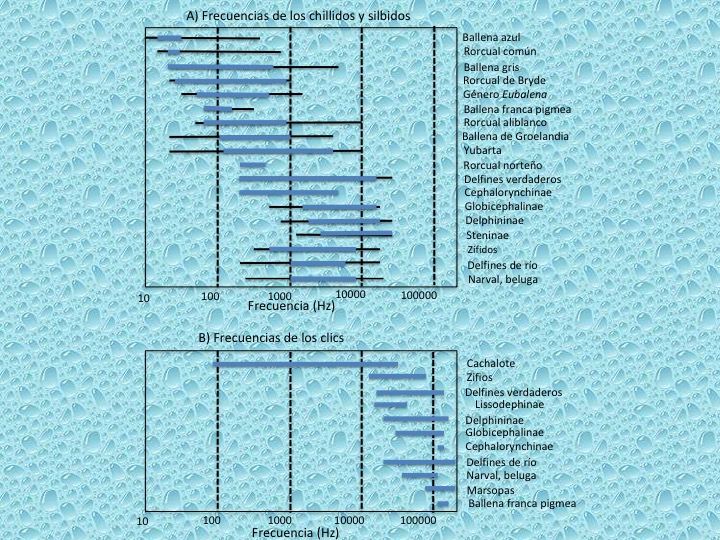
Los misticetos no hacen uso de la ecolocalización. Carecen de canal y grasa mandibular. Ellos producen sonidos de alto volumen en el rango infrasónico. Se trata de sonidos de amplitud modulada que viajan a mayores distancias que los ultrasonidos.
El comportamiento de vocalización varía con el género, edad y temporada. Por ejemplo, adultos machos de muchas especies de misticetos vocalizan regularmente y en voz alta durante la temporada de cría. Los sonidos bajos viajan particularmente bien en el mar, perdiendo energía por conversión a calor sólo después de recorrer distancias muy largas. Así, las intensas vocalizaciones de algunas especies se pueden detectar a distancias de varias decenas de kilómetros.
La ecolocalización en los cetáceos:
En los cetáceos odontocetos (delfines, orcas, calderones, marsopas, cachalotes, etc.) una estructura, el melón, rellena de espermaceti o "esperma de ballena" (sustancia de naturaleza lipídica), situada en la frente de estos animales y ausente en las llamadas "ballenas con barbas", interviene en la ecolocalización proyectando y dirigiendo las ondas producidas hacia el frente.
La ecolocalización consiste en la emisión de sonidos de alta frecuencia y la capacidad de detectar su eco, después de reflejarse en objetos y obstáculos. Las ondas sonoras se forman en los sacos nasales del animal, pasan al melón, el cual actúa como una lente acústica. El melón concentra las ondas sonoras y las transforma en un haz que se proyecta hacia adelante. Este haz, como ondas acústicas, viaja a través del agua y rebotan al tropezar con los objetos regresando al cetáceo en forma de eco (Figura 16). Una vez recibidos a través de los canales de grasa de la mandíbula inferior, esos ecos son analizados y conducidos hasta el oído medio (hueso timpánico-periótico) hasta llegar al cerebro (Figura 17). De esta forma son capaces de determinar el tamaño, la forma, la estructura, la composición, la velocidad y dirección de los mismos. Las altas frecuencias se disipan antes que frecuencias más bajas, pero permiten detectar estructuras más pequeñas y mayores detalles. Son capaces de detectar objetos a más de 70 metros y hay evidencias que sugieren que podrían variar las frecuencias de sus clics dependiendo del ambiente y del objeto a ecolocalizar para no competir con el ruido de fondo.
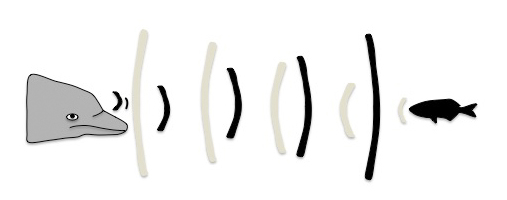
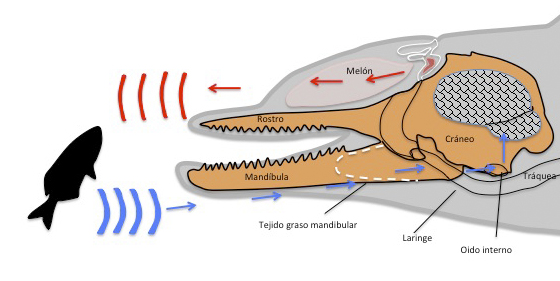
Los odontocetos producen los sonidos en la región nasal gracias a los sacos aéreos. Carecen de cuerdas vocales. Todos ellos cuentan con una par de estructuras conocidas como “labios fónicos” (monkey lips) que se encuentran conectados a dos sacos llenos de lípidos que se llaman bursas dorsales. Conjuntamente las bursas dorsales (anterior y posterior) y los labios fónicos forman el complejo MLDB que parece constituir la estructura generadora de los sonidos (Figura 18). El sonido se crea cuando el aire pasa a través de los labios fónicos y produce vibraciones y el aire utilizado para crearlo puede volver al canal nasal o ser liberado al agua (burbujas). La vibración producida es transmitida al melón, estructura rica en lípidos, que focaliza el sonido en el agua.
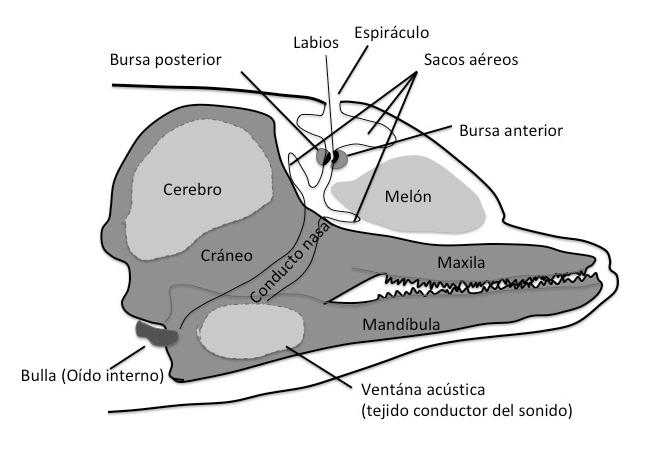
El cerebro de los delfines no cuenta con un área olfativa definida ni receptores químicos en sus tejidos respiratorios. Tampoco se ha descrito de una manera clara un nervio olfativo. Se piensa que el sentido del olfato se perdió al transformarse los conductos nasales en generadores de sonido.
El sentido del gusto parece estar presente, al menos, en algunas especies de cetáceos. Varias especies de odontocetos presentan en la lengua lo que parecen ser papilas gustativas. Se ha comprobado que los delfines mulares (Tursiops truncatus), pueden detectar los cuatro gustos primarios (dulce, salado, ácido y amargo), y en ambientes controlados muestran preferencias de comida. A pesar de esto, se cree que el gusto es importante en el reconocimiento de las diferentes secreciones corporales utilizadas para la señalización, como pueden ser la receptividad sexual o el peligro.
Cada una de las especies de cetáceos ha desarrollado una combinación única de estrategias de comportamientos así como adaptaciones fisiológicas para asegurar la supervivencia.
La adaptación de los cetáceos a la vida acuática y la adquisición de la forma hidrodinámica del cuerpo, ha conllevado también modificaciones en los genitales externos. En la mayoría de las especies es difícil distinguir los sexos a primera vista porque el pene del macho queda retraído en el prepucio (excepto cuando está erecto), cuya abertura es un pliegue genital de aspecto parecido al pliegue genital de la hembra o vulva. La única diferencia visible entre ambos sexos es la distancia que hay entre el ano y el pliegue genital, ésta en el macho es aproximadamente el 10% de la longitud corporal, pero en la hembra, el ano y el pliegue genital parecer estar unidos. Incluso las glándulas mamarias son difíciles de distinguir, ya que los pezones que están uno a cada lado del pliegue genital, quedan dentro de un pequeño pliegue mamario (Figura 19). El pezón sobresale únicamente durante la lactancia.
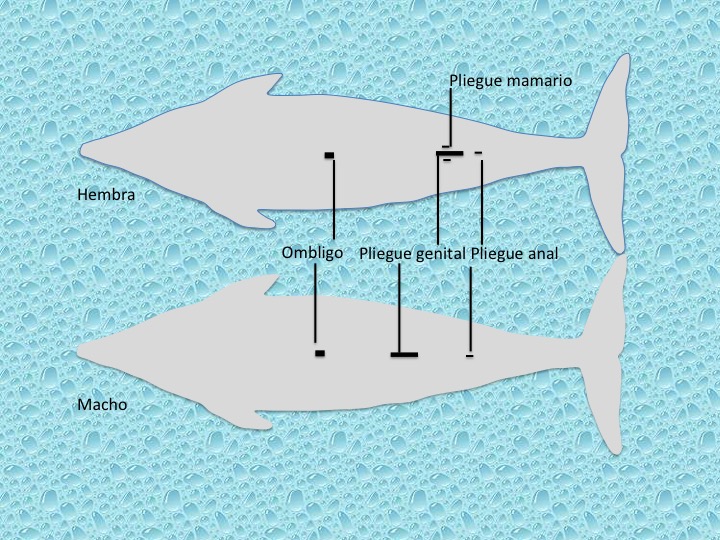
Existe dimorfismo sexual en muchas especies de cetáceos, siendo más evidentes en unas que en otras. Este dimorfismo sexual puede manifestarse en el tamaño del cuerpo, como es el caso del cachalote (Physeter macrocephalus), en el tamaño y forma de las aletas dorsales como es el caso de la orca (Orcinus orca), así como en la adquisición de ciertos caracteres en uno de los sexos, como es el caso del narval (Monodon monoceros). También pueden aparecer diferentes tonalidades, dentro de las coloraciones características de una especie, entre el macho y la hembra como le ocurre al delfín rosado (Inia geoffrensis).
CLASIFICACIÓN DE LOS CETÁCEOS
El orden Cetacea incluye actualmente casi 90 especies agrupadas en 42 géneros, 14 familias y dos subórdenes: Odontocetos (cetáceos con dientes) y Misticetos (cetáceos con barbas).
TAXONOMÍA DE ODONTOCETOS: cetáceos con dientes


El Mar de Alborán, a pesar de ser una de las rutas de transporte marítimo más transitadas del mundo, es uno de los puntos con mayor biodiversidad del mar Mediterráneo. En él se pueden observar varias especies de cetáceos, tanto de odontocetos como de misticetos.
Entre ellas, hay algunas que pueden observarse prácticamente todo el año, por eso se le denominan Especies Residentes (aunque no está demostrado que realmente estén todos los días del año) y son 4 especies de odontocetos: el delfín listado (Stenella coeruleoalba, Meyen, 1833), el delfín común (Delphinus delphis, Linnaeus, 1758), delfín mular (Tursiops truncatus, Montagu, 1821) y calderón común de aleta larga (Globicephala melas, Traill, 1809).
Delfín común (Delphinus delphis) cazando en la Bahía de la Herradura
Las especies denominadas Semi-residentes, se pueden observar en determinados periodos del año en los que la comida es más abundante. Dentro de esta categoría, estarían otras dos especies de odontocetos, la orca (Orcinus orca, Linnaes, 1758 y el cachalote (Physeter macrocephalus, Linnaeus, 1758). La primera se observa entre los meses de julio a septiembre y la segunda entre marzo y julio con mayor probabilidad. Por último, hay una especie de misticetos, el Rorcual común (Balaenoptera physalus, Linnaeus, 1758), que pasa por el Estrecho de Gibraltar en su ruta migratoria. Los meses en los que hay mayor probabilidad de avistarlos va desde mayo a julio.
 |
 |
La navegación, contaminación, redes de pesca, etc., suponen un peligro para estas especies. En las imágenes siguientes se puede observar un ejemplar de Megaptera novaeangliae, que previamente fue rescatado de una red en Mallorca, varado en una playa de Valencia el 26 de mayo de 2022.
 |
 |
 |
 |
 |
BIBLIOGRAFÍA
- Alan N. Baker, Whales and dolphins of New Zealand and Australia: an identification guide. Wellington: Victoria University Press, 1999, pp. 42–43.
- Au, W. W. L. (1993). The Sonar of Dolphins. Springer, New York.
- Berta, A., Sumich, J. L., & Kovacs, K. M. (2005). Marine mammals: evolutionary biology. Academic Press.
- Bostrom, B. L., Fahlman, A., & Jones, D. R. (2008). Tracheal compression delays alveolar collapse during deep diving in marine mammals. Respiratory physiology & neurobiology, 161(3), 298-305.
- Carwardine, M. (1998). Manual de Identificación Ballenas. Delfines y Marsopas: Guía Visual de todos los cetáceos del mundo.
- Fahlman, A., Tyack, P. L., Miller, P. J., & Kvadsheim, P. H. (2014). How man-made interference might cause gas bubble emboli in deep diving whales. Frontiers in physiology, 5.
- Hicks, B. D., Aubin, D. J. S., Geraci, J. R., & Brown, W. R. (1985). Epidermal growth in the bottlenose dolphin, Tursiops truncatus. Journal of Investigative Dermatology, 85(1), 60-63.
- Hill, R. W., Wyse, G. A., Anderson, M., & Anderson, M. (2004). Animal physiology (Vol. 2). Massachusetts: Sinauer Associates.
- Hooker, S.K. (2009). Encyclopedia of Marine Mammals.
- Kinze, C.C. (2002). Mamíferos Marinos del Atlántico y del Mediterráneo. Ed. Omega.
- Kooyman, G. L. (1973). Respiratory adaptations in marine mammals. American Zoologist, 13(2), 457-468.
- Martin, A. R., & Reeves, R. R. (2002). Diversity and zoogeography. Marine mammal biology: An evolutionary approach, 1-37.
- Mellinger, D. K., Stafford, K. M., Moore, S. E., Dziak, R. P., & Matsumoto, H. (2007). An overview of fixed passive acoustic observation methods for cetaceans. Oceanography, 20(4), 36-45.
- Ollivier, F. J., Samuelson, D. A., Brooks, D. E., Lewis, P. A., Kallberg, M. E., & Komáromy, A. M. (2004). Comparative morphology of the tapetum lucidum (among selected species). Veterinary ophthalmology, 7(1), 11-22.
- Perrin, W. F., Würsig, B., & Thewissen, J. G. M. (Eds.). (2009). Encyclopedia of marine mammals. Academic Press.
- Ridgway, S. H. (1986). Diving by cetaceans. Diving in animals and man. The Royal Norwegian Society of Science and Letters, Trondheim, Norway, 33-62.
- Watkins, W. A. (1968). The harmonic interval: fact or artifact in spectral analysis of pulse trains. Woods Hole Oceanographic Institution.
LILIANA OLAYA PONZONE