
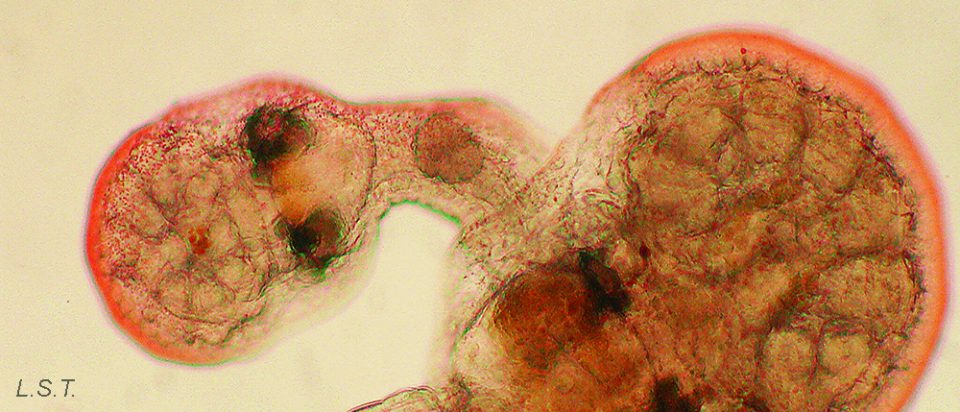
Reino: Animal
Filo ENTOPROCTA
El filo endoproctos (del griego ἐνδο, 'dentro' + πρωκτός, 'ano') (= kampozoos) está compuesto por cuatro familias actualmente aceptadas: Barentsiidae Emschermann, 1972, Loxokalypodidae Emschermann, 1972, Loxosomatidae Hincks, 1880 y Pedicellinidae Johnston, 1847. En total incluye cerca de 200 especies agrupadas en unos 12 géneros. No obstante, se trata de un grupo muy poco estudiado y, como señaló el Dr. C. Nielsen, máxima autoridad mundial en el conocimiento de este filo, el número real de especies que lo componen podría ser de unas 500. Fósiles de este grupo animal datan del Jurásico superior.
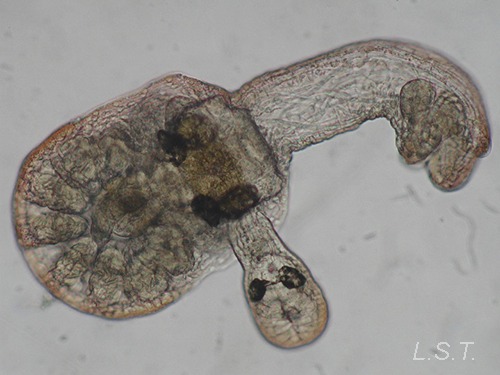
Hábitat y distribución
Todas las especies de endoproctos conocidas viven en medios marinos o salobres a excepción de dos de ellas: Urnatella gracilis (familia Barentsiidae) y Loxosomatoides sirindhornae (familia Pedicillinidae). Mientras la última, recientemente descrita, solo ha sido citada en Tailandia, U. gracilis presenta una distribución casi cosmopolita. Las restantes especies del grupo se distribuyen por áreas marinas de todo el planeta, especialmente en zonas poco profundas (0-500 metros).
Viven fijos a substratos de distinta naturaleza (aunque, como se tratará más adelante, algunos son capaces de desplazarse activamente), tanto abiótica como biótica (rocas, conchas, otros invertebrados) y son frecuentes los casos de comensalismo sobre otras especies marinas de vida sésil como esponjas, briozoos, poliquetos, ascidias, etc.

Los endoproctos pueden ser solitarios (como ocurre en todos los loxosomátidos) o coloniales (los miembros de las restantes familias).
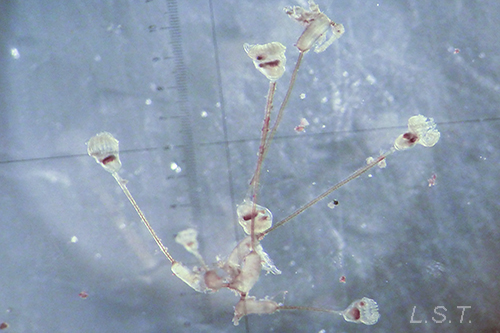
Morfología
Los endoproctos son pequeños animales que raras veces superan 1 mm de longitud total (aunque su tamaño oscila entre 0,1 y 7 mm). Su cuerpo está formado por un cáliz, en cuyo interior se encuentran las vísceras, y un pedúnculo que parte de él y que en su extremo distal se ancla al substrato. En las formas coloniales, los zooides están interconectados por sus pedúnculos que pueden adoptar formas ramificadas o partir de un estolón común. Los pedúnculos poseen musculatura.
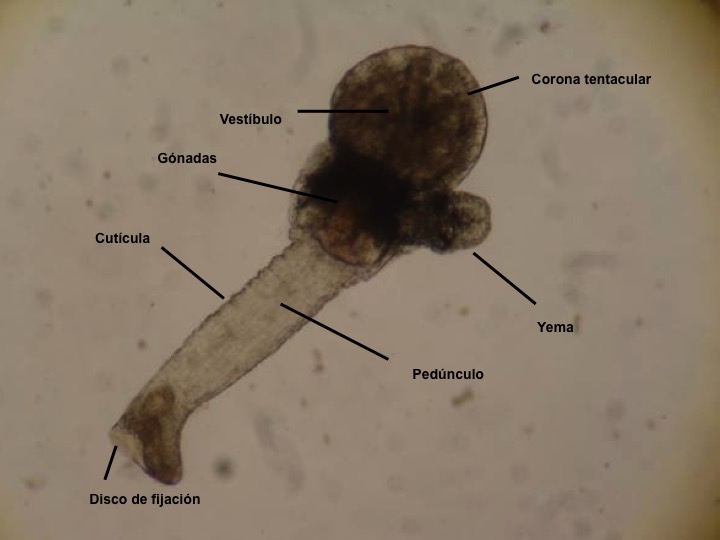
El cáliz porta una corona de tentáculos constituida por entre 8 y 30 unidades que rodean un atrio o vestíbulo donde se hallan la boca y el ano (así como el/los nefridioporo/s y el gonoporo).
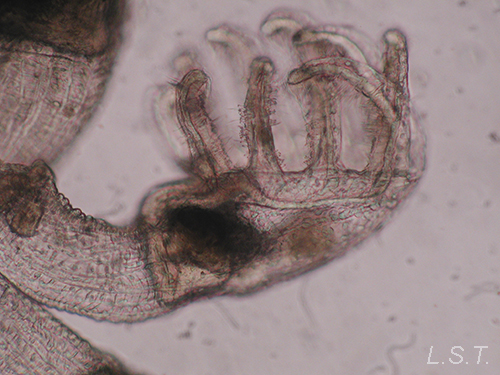
La base de los tentáculos está recubierta por una membrana que se repliega sobre el atrio cuando los tentáculos están retraídos. Las estructuras sensoriales se disponen en los tentáculos. El pedúnculo posee musculatura longitudinal y también aparece musculatura asociada a los tentáculos, la membrana y algunas áreas del cáliz. El cáliz y el pedúnculo están cubiertos por una fina cutícula, reducida en la parte ciliada de los tentáculos y en el vestíbulo. El tubo digestivo tiene forma de U e incluye un estómago voluminoso.
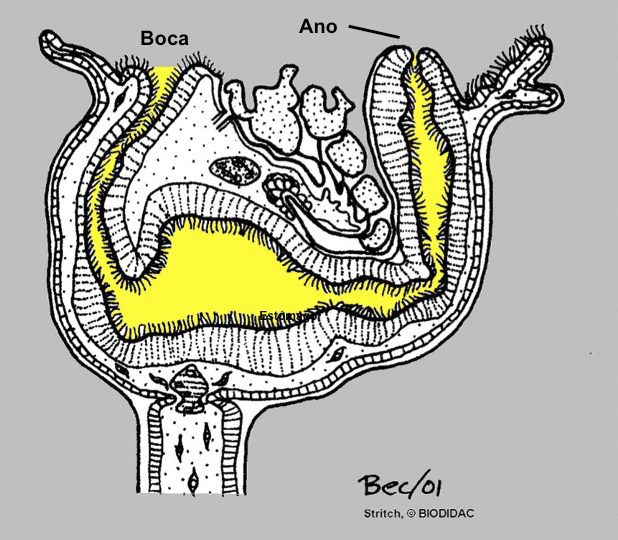
El sistema excretor está formado por protonefridios y el sistema nervioso es subepidérmico con un ganglio con forma de pesa (y del que parten los nervios) próximo al estómago, junto al cual también están las gónadas (uno o dos pares). Carecen de sistema respiratorio diferenciado, pues el intercambio de gases tiene lugar a través de la superficie corporal, particularmente en las superficies más finas. Las especies de las familias Pedicellinidae y Barentsiidae poseen un complejo de células con forma de estrella en la zona de transición entre el cáliz y el pedúnculo que está ausente en las restantes dos familias.
Alimentación
Los endoproctos ingieren partículas orgánicas del medio junto con algas, bacterias y otros microorganismos del plancton. Aunque son capaces de generar corrientes de agua, las especies comensales con frecuencia se ven beneficiadas del flujo generado por su hospedador cuando éste es un animal filtrador como una esponja o una ascidia.
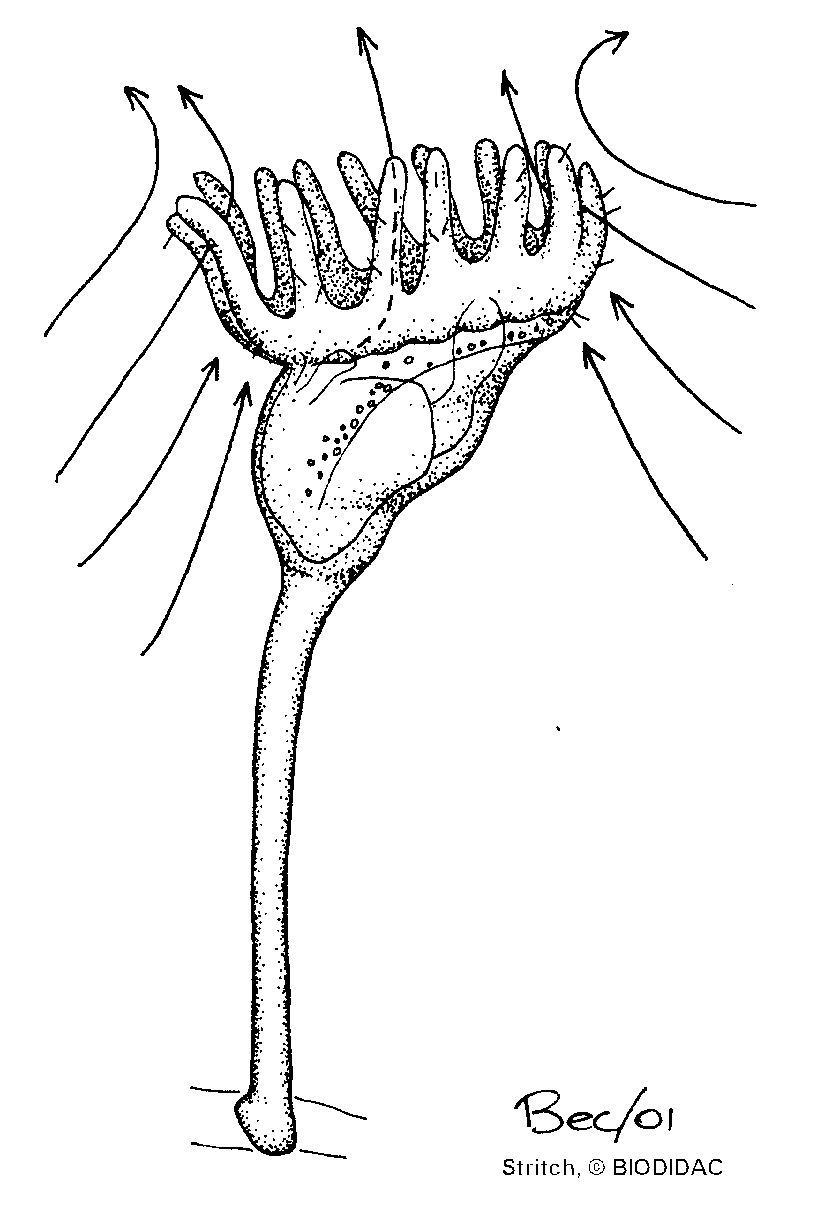
Para producir las corrientes de agua los endoproctos baten los cilios laterales de los tentáculos, lo que hace que el agua pase hacia el atrio dejando las partículas de alimento retenidas en los tentáculos.
Los cilios frontales de los tentáculos son los responsables de transportar dichas partículas hasta la base de los tentáculos y, desde allí, llegan a la boca por unos surcos vestibulares que también están ciliados. Las glándulas de la corona de tentáculos producen secreciones pegajosas que ayudan a capturar las partículas.
Reproducción
Los endoproctos son animales que se reproducen tanto sexual como asexualmente.
La reproducción asexual se produce por gemación. En las especies solitarias las yemas formadas en el cáliz se desprenden y se fijan al substrato, mientras que en las especies coloniales las yemas suelen salir del estolón o las ramas de la colonia incrementando el tamaño de la misma.
En la reproducción sexual, la fecundación parece ocurrir en el interior del ovario o del oviducto, aunque el conocimiento en este aspecto es muy escaso. En las formas solitarias los adultos parecen ser hermafroditas protándricos, mientras que en las formas coloniales se han encontrado especies con colonias unisexuales (machos o hembras), otras con colonias que incluyen zooides machos y zooides hembras, y otras con zooides hermafroditas simultáneos. En muchas especies los huevos se incuban externamente en el vestíbulo. Del huevo nace la larva (de tipo trocófora, como la de anélidos y moluscos) que llevará una vida libre (que dura de unas pocas horas a varias semanas según la especie) hasta su fijación y metamorfosis para originar el adulto directamente o a través de la producción de yemas. Aunque predominan las larvas de vida libre planctotrófica, algunas especies producen larvas lecitotróficas o larvas bentónicas. Existen también especies vivíparas en las que el embrión se desarrolla en el ovario y es nutrido por su progenitor.
Relaciones filogenéticas
Los endoproctos son considerados como un grupo monofilético. Las relaciones de parentesco evolutivo de este grupo con otros filos animales continúan siendo muy discutidas. Si bien se suele aceptar una proximidad con el filo ciclióforos (apoyado tanto por estudios de ultraestructura como moleculares), más discutida es su relación con los ectoproctos (= briozoos). La presencia de una larva tipo trocófora los sitúa sin duda dentro del gran grupo de los lofotrocozoos.
Depredadores
Uno de los depredadores más especializados en la captura de endoproctos son los nudibranquios del género Trapania. Éstos poseen una rádula que consta de dos filas de dientes, compuestos a su vez por numerosos dentículos puntiagudos, que actúan a modo de peine, arrancando a los endoproctos por su base e ingiriéndolos enteros.

Endoproctos en el litoral granadino
En las costas de Granada tan solo se han citado representantes de la familia loxosomátidos, que se caracteriza por que el cáliz y el pedúnculo no aparecen separados por un septo y los músculos pasan entre ambos sin interrupción, carecen del complejo de células con forma de estrella, presentan un cono anal, los protonefridios tienen nefridioporos separados y por ser los únicos endoproctos solitarios (no coloniales). Los Loxosomatidae son generalmente comensales de otros invertebrados marinos como esponjas, ascidias, sipuncúlidos, ectorpoctos o poliquetos y cada especie suelen presentar especificidad en cuanto a su hospedador, de manera que cada especie de endoprocto se asocia a una o pocas especies de hospedadores. Aunque viven unidos al substrato, algunos pueden desplazarse arrastrándose y volteándose.
Los Loxosomátidos constituyen la familia más diversa dentro de los endoproctos al incluir tres cuartas partes de todas las especies de este filo. Dentro de esta familia, el género Loxosomella es el más diversificado, con unas 110 especies descritas. Las yemas de Loxosomella presentan un característico pie con una glándula conectada a un surco que está bordeado de una fila de conspicuas células y, así mismo, pueden presentar estructuras unicelulares de fijación en el extremo del pie o en la fila de células que bordea el surco. Los adultos pueden conservar estas estructuras de anclaje o degenerarlas, en cuyo caso los individuos se convierten en organismos fijos al substrato e incapaces de desplazarse.
La costa granadina ha sido bien estudiada en lo que respecta a su fauna de endoproctos y cinco especies de Loxosomella han sido citadas en esta área, dos de ellas descritas como especies nuevas a partir de colectas realizadas en la costa de Almuñécar: L. almugnecarensis Tierno de Figueroa & Sánchez-Tocino, 2009 y L. ameliae Sánchez-Tocino & Tierno de Figueroa, 2009. Las otras tres especies citadas son L. crassicauda (Salensky, 1877), L. pes (Schmidt, 1878) y L. tethyae (Salensky, 1877). Algunas de estas especies pueden formar poblaciones muy numerosas sobre sus hospedadores, especialmente sobre las esponjas, pero aun así, debido a su pequeño tamaño, no son fácilmente detectables. En nuestra costa, sí que pueden indicar su presencia la aparición de nudibranquios del género Trapania Pruvot-Fol, 1931 alimentándose sobre esponjas de los géneros Ircinia Nardo, 1833 y Sarcotragus Schmidt, 1862, ya que, como hemos comentado, estas babosas de mar son activos depredadores de los endoproctos.
- Family Loxosomatidae Hincks, 1880
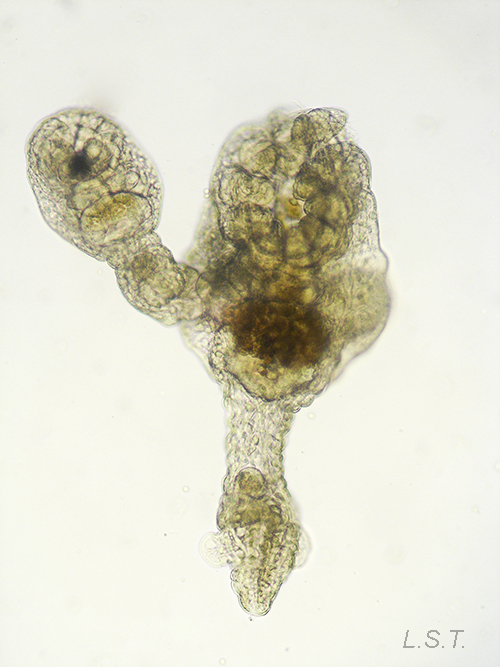
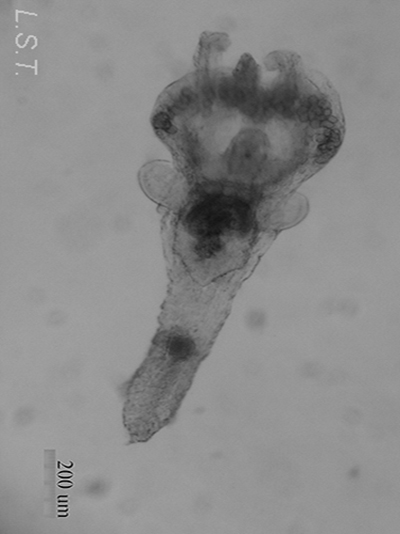
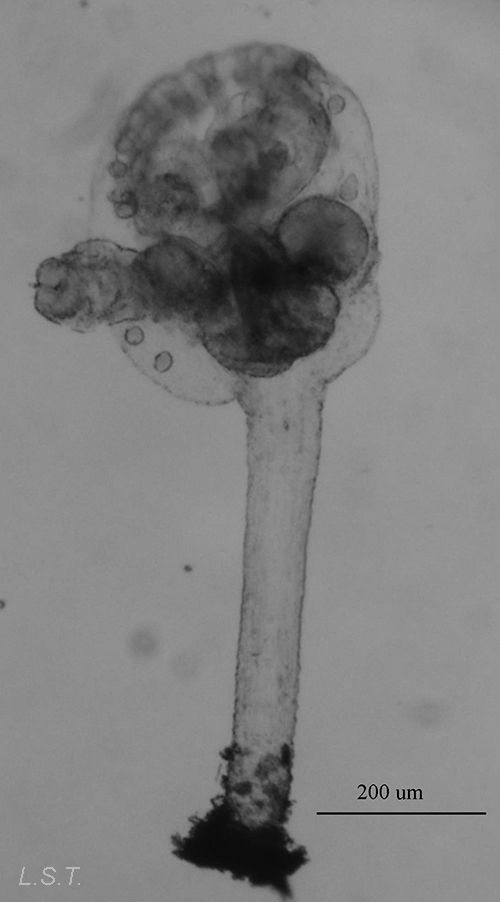
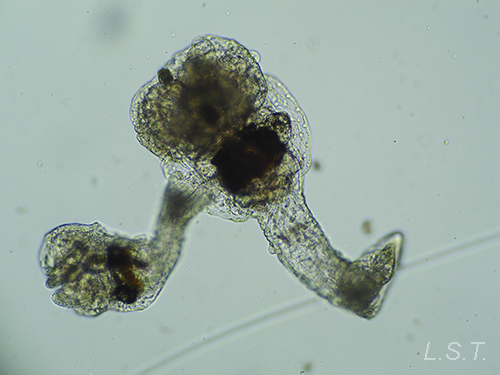
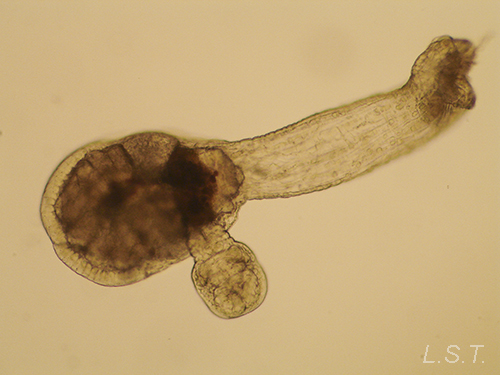
- Familia PEDICELLINIDAE

ESPECIES SIN IDENTIFICAR
|
|
|


|
|
|


|
|

02-05-2025 Ensenada de los Gigantes -16m
MÁS INFORMACIÓN
VÍDEOS
07-11-2020 Punta de la Mona -11 m.
BIBLIOGRAFÍA
Litoral granadino
- SÁNCHEZ-TOCINO, L. y J.M. TIERNO DE FIGUEROA. 2009. Contribution to the knowledge of Loxosomatidae (Entoprocta) from the Spanish Mediterranean Sea, with the description of a new species: Loxosomella ameliae sp.n. Marine Biology Research, 5: 404-407.
- SÁNCHEZ-TOCINO, L. y J.M. TIERNO DE FIGUEROA. 2009. Los endoproctos del Mar de Alborán. Quercus, 284: 42-44.
- TIERNO DE FIGUEROA, J.M. y L. SÁNCHEZ-TOCINO. 2009. Loxosomella almugnecarensis n. sp. (Entoprocta: Loxosomatidae) ― a new sponge epizoite from the Iberian Mediterranean Sea. Zootaxa, 2236: 65-68.
- TIERNO DE FIGUEROA, J.M. y L. SÁNCHEZ-TOCINO. 2008. First record of Loxosomella crassicauda (Salensky, 1877) (Entoprocta, Loxosomatidae) in the Mediterranean Spanish waters. Zoologica baetica, 19: 89-93.
- TIERNO DE FIGUEROA, J.M. y L. SÁNCHEZ-TOCINO. 2009. First record of Loxosomella tethyae (Salensky, 1877) (Endoprocta, Loxosomellidae) in the Iberian Mediterranean coast. Zoologica baetica, 20: 101-104.
General
- Brusca, R.C. y Brusca, G.J. 2003. Invertebrates. 2nd edition. Sianauer Associates, Inc., Publishers, Sunderlands, Massachusetts. 922 pp.
- Emschermann, P. 1982. Les Kamptozoaires. État actuel de nos connaissances sur leur anatomie leur Développement, leur biologie et leur position phylogénétique. Bulletin delle Société Zoologique de France, 107 (2): 317-344.
- NIELSEN, C. 1989. Entoprocta. Synopses of the British Fauna, n.s., 41: 1-131.
- NIELSEN, C. 2010. A review of the taxa of solitary entoprocts (Loxosomatidae). Zootaxa, 2395: 45-56.
- Nielsen, C. 2012. Animal evolution. Interrelationships of the living phyla. Third edition. Oxford University Press, Oxford. 402 pp.
- Ruppert, E.E., R.S. Fox y R.D. Barnes. (2004). Invertebrate Zoology. A functional Evolutionary Approach. 7th edition. Brooks/Cole, Belmont. 1008 pp.
- Sánchez-Tocino, L. y J.M. Tierno de Figueroa. 2009. Contribution to the knowledge of Loxosomatidae (Entoprocta) from the Chafarinas Islands (Alboran Sea, Western Mediterranean). Graellsia, 65(1): 71-74.
- Westheide, W. y Rieger, R. 2011. Zoologia sistematica. Filogenesi e diversità degli animali. (Edizione italiana a cura di A. Minelli). Zanichelli editore, Bologna. 1048 pp.
- WOOD, T.S. 2005. Loxosomatoides sirindhornae, new species, a freshwater kamptozoan from Thailand (Entoprocta). Hydrobiologia, 544: 27-31.
- Yakovis, E.L. 2002. Substrate preferences of a non-colonial kamptozoan, and its interactions with bryozoan hosts. Marine Biology, 141: 1109-1115.
PÁGINAS WEB
- WORMS (http://marinespecies.org/aphia.php?p=taxdetails&id=1271)
JOSÉ MANUEL TIERNO DE FIGUEROA